Классификация белков.
Биологические функции белков

Эти остатки оказываются включенными в белок не в ходе его матричного синтеза, а в результате химического посттрансляционного превращения входящих в его состав аминокислот. Гидроксилирование пролина требует в качестве кофактора (небелкового компонента, необходимого для эффективной работы) аскорбиновую кислоту (витамин С), которая нужна для поддержания в восстановленном состоянии иона Fe2… Читать ещё >
Классификация белков. Биологические функции белков (реферат, курсовая, диплом, контрольная)
Сложность строения белковых молекул и чрезвычайное разнообразие их функций крайне затрудняют создание единой четкой их классификации на какой-либо одной основе. Белки можно классифицировать по их составу (простые, сложные), структуре (фибриллярные, глобулярные, промежуточные), функциям. Рассмотрим подробнее структурную классификацию.
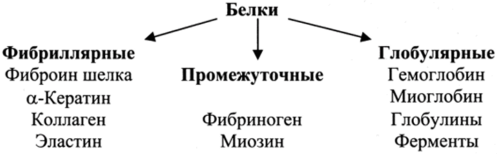
Фибриллярные белки сильно вытянуты (наиболее важна вторичная структура) и выполняют структурные функции.
Глобулярные белки, которые в грубом приближении могут быть представлены в виде сфер (наиболее важной является третичная структура), принимают участие в таких специфических процессах, как катализ, транспорт, регуляция.
Кроме перечисленных выше типов белков, в организме имеются небольшие или бедные углеводородными группами полипептиды, которые могут сами по себе не иметь фиксированной структуры, но приобретать ее при взаимодействии с другими макромолекулами. Следует отметить, что данная классификация не может претендовать на полноту, так как существуют белки, которые не относятся ни к одному из этих классов. Например, миозин, который по своей структуре содержит признаки и фибриллярного и глобулярного белка.
Белок с исходной, природной укладкой цепи, т. е. имеющий трехмерную конфигурацию, называется нативным, белок с развернутой, беспорядочной укладкой цепи — денатурированньш. Превращение нативного белка в денатурированный, т. е. утрата белком его трехмерной конфигурации, называется денатурацией (рис. 3.15). Вызывать денатурацию могут разнообразные факторы. В частности, плотная укладка цепи белка обычно нарушается при нагревании. Тепловая денатурация — общее свойство белков. После денатурации биологически активный белок может самопроизвольно свернуться в исходную конформацию с восстановлением своей активности. Процесс сворачивания денатурированного белка называется ренатурацией.
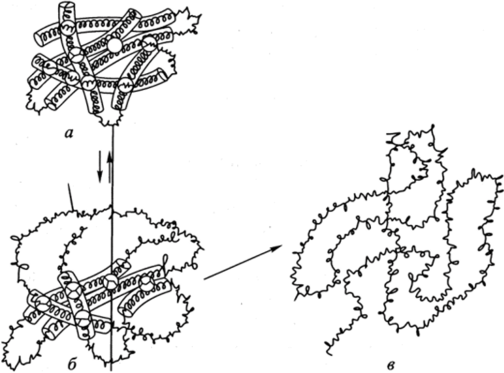
Рис. 3.15. Денатурация белковой молекулы:
а — исходное состояние; б — начинающееся обратимое нарушение молекулярной структуры; в — необратимое развертывание полипептидной цепи При длительном воздействии денатурирующего агента (температуры, химического вещества, среды с различным pH) денатурация становится необратимой (на рис. 3.15 этот процесс обозначен стрелкой между состояниями белковой молекулы б и в). Большинство белков денатурирует при нагревании их растворов выше 50−60 °С.
Денатурированный белок теряет способность растворяться в воде. Наиболее характерным признаком денатурации является резкое снижение или полная потеря белком его биологической активности (каталитической, антигенной или гормональной). Тот факт, что денатурированный белок полностью теряет свои биологические свойства, подтверждает тесную связь между структурой белковой молекулы и функцией, которую она выполняет в организме.
Способность белковой молекулы спонтанно ренатурироваться при снятии внешнего агрессивного воздействия говорит о том, что аминокислотная последовательность сама определяет пространственную структуру белка без участия какого-либо внешнего регулирующего центра.
В настоящее время денатурация и ренатурация глобулярных белков in vitro интенсивно исследуются, так как эти процессы связаны с проблемой самоорганизации белка, т. е. с вопросом о том, как белковая цепь «находит» свою уникальную структуру среди гигантского числа возможных альтернатив.
Фибриллярные белки составляют основу не растворимых в воде и прочных материалов, таких как рога, копыта, ногти, шерсть, волосы, перья, кожа, сухожилия, межклеточное вещество костной ткани. Волос — длинное достаточно прочное волокно, основой которого является белок — а-кератин. В основе сухожилий другой белок — коллаген. Эластичность и упругость стенкам артерий или легочных альвеол придает эластин. Общей особенностью этих белков является участие в формировании их пространственной структуры ковалентных непептидных связей.
Кератины волос и шерсти образуют промежуточные филаменты, состоящие из длинных полипептидных цепей с крупными доменами, образованными а-спиралями и содержащими повторяющиеся последовательности из семи аминокислотных остатков (гептапептиды). Две направленные одинаково цепи кератина образуют суперспираль, в которой остатки неполярных аминокислот обращены внутрь и тем самым защищены от воздействия воды. Такая структура дополнительно стабилизируется многочисленными дисульфидными связями, образованными остатками цистеина соседних цепей. Суперспиральные димеры, в свою очередь, объединяются с образованием тетрамеров, подобных четырехжильному канату.
Коллаген образуется вне клеток из секретируемого ими белка — проколлагена, который превращается в коллаген в результате взаимодействия соответствующих ферментов. Молекула проколлагена представляет собой тройную суперспираль, образованную тремя скрученными вместе специализированными полипептидами. Далее при отщеплении концевых полипептидов образуется тропоколлаген, который упаковывается в коллагеновые волокна. Каждый из трех полипептидов в тропоколлагене находится в виде левосторонней спирали (в отличие от обычных правосторонних а-спиралей у белков). Примерно треть аминокислотных остатков в тропоколлагене представлена пролином, а каждый третий остаток — глицином.
В ходе образования коллагена многие остатки пролина и лизина в присутствии аскорбиновой кислоты гидроксилируются, превращаясь соответственно в гидроксипролин и гидроксилизин:

Эти остатки оказываются включенными в белок не в ходе его матричного синтеза, а в результате химического посттрансляционного превращения входящих в его состав аминокислот. Гидроксилирование пролина требует в качестве кофактора (небелкового компонента, необходимого для эффективной работы) аскорбиновую кислоту (витамин С), которая нужна для поддержания в восстановленном состоянии иона Fe2+ в активном центре фермента прол ил-гидроксил азы. При недостатке витамина С нарушается образование соединительных тканей, что вызывает тяжелое заболевание — цингу.
Три спирально навитые друг на друга молекулы тропоколлагена ковалентно связаны между собой, образуя прочную структуру. Такая ассоциация невозможна в обычной белковой спирали, так как этому препятствуют объемные боковые цепи. В коллагене спирали более вытянуты (на один виток приходится 3 остатка, вместо 3,6), так как каждый третий аминокислотный остаток — глицин, поэтому спирали в этих точках максимально приближены друг к другу. Дополнительная стабилизация структуры осуществляется водородными связями гидроксилированных остатков лизина и пролина.
Молекулы тропоколлагена содержат около 1000 аминокислотных остатков. Они собираются в коллагеновые фибриллы, стыкуясь «голова к хвосту». Пустоты в этой структуре при необходимости могут служить местом первоначального отложения кристаллов гидроксиапатита Са5(0Н)(Р04)з, играющего важную роль в минерализации костей.
Коллаген сухожилий подвергается ферментативной модификации — в концевых частях тропоколлагеновых цепей ковалентно сшиваются остатки лизина. Таким образом, сухожилия представляют собой пучки параллельно ориентированных фибрилл. В отличие от сухожилий в коже коллагеновые фибриллы образуют подобие неупорядоченной двумерной сетки.
Эластин по своему строению отличается от коллагена и акератина. Он содержит обычные а-спирали, образующие поперечно-сшитую сеть, которая своей необычайно высокой эластичностью обязана уникальному способу связывания боковых цепей лизина:
четыре сближенных лизиновых остатка 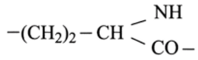
формируют так называемую десмозиновую структуру, объединяющую в один узел четыре участка пептидных цепей (рис. 3.16).
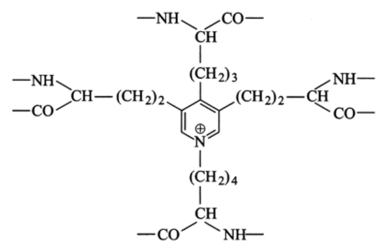
Рис. 3.16. Химическая структура десмозина.
Глобулярные белки. Большинство белковых молекул в организме имеет глобулярное строение. Пептидная связь в глобулярных белках в естественном состоянии свернута в компактные структуры — глобулы, которые в первом грубом приближении могут быть представлены в виде шара или не слишком вытянутого эллипсоида, в отличие от фибриллярных белков, где длинные полипептидные цепи вытянуты вдоль одной оси.
Глобулы устойчивы в водных системах вследствие того, что полярные группы основной и боковых цепей сосредоточены на поверхности, находясь в контакте с водой, а неполярные обращены в глубь молекулы и защищены от этого контакта. На поверхности белковой глобулы иногда образуются ионные связи — солевые мостики.
Оказавшиеся внутри глобулы >N-H и >С=0-группы основной цепи с образовавшимися водородными связями формируют в результате а-спирали и (3-слои. Дестабилизирующим фактором пространственной упаковки является наличие в глубине глобулы каких-то групп, потенциально способных образовывать ионные и водородные связи, но реально лишенных партнеров.
При физиологических условиях состояние белка, имеющего нативную трехмерную структуру, термодинамически стабильно, т. е. соответствует минимуму свободной энергии. Информация, необходимая для сворачивания белка в нативную конформацию, заложена в его аминокислотной последовательности. Поэтому в принципе теоретически можно предсказать трехмерную структуру любого белка исходя из его аминокислотной последовательности. Однако предсказание третичной структуры остается нерешенной проблемой молекулярной биологии. Сворачивание молекулы белка из развернутого состояния должно осуществляться единственным путем. Если предположить, что белковая молекула состоит из 50 остатков, каждый из которых может принимать 10 разных конформаций, то общее число возможных конформаций составит 1050, и если характерное время молекулярных перестроек составляет 10″13 с, то для того, чтобы перепробовать все конформации, потребуется 1037 с (~ Ю30 лет). Следовательно, существует направленный путь сворачивания белка.
Стабильность свернутой молекулы белка в водном окружении крайне низка. Основной движущей силой сворачивания является энтропийный гидрофобный эффект, вследствие которого неполярные группы стремятся выйти из водного окружения и оказаться внутри глобулы. Существует и обратный эффект, препятствующий сворачиванию и обусловленный тем, что для свернутой молекулы белка число разрешенных конформаций основной и боковых цепей меньше, чем у развернутой.
Гемоглобин (НЪ) — белок, переносящий кислород от легких к тканям. НЬ локализован в красных кровяных клетках — эритроцитах.
Как уже отмечалось (см. рис. 3.14), гемоглобин состоит из четырех полипептидных цепей, каждая из которых содержит гем (рис. 3.17). Функциональная взаимосвязь этих цепей такова, что присоединение О2 к одному из атомов железа повышает сродство к кислороду у трех других.
Гемоглобины — это целый класс белков, представители которого различаются одним-двумя аминокислотными остатками или их последовательностью. У взрослого человека гемоглобин типа НЬА. Кроме НЬА, существует эмбриональный гемоглобин HbF, исчезающий после рождения. Молекулярная масса обоих гемоглобинов приблизительно одинакова (64 500), они отличаются только последовательностью аминокислотных остатков. Наряду с обычно имеющимися гемоглобинами в организме человека встречаются аномальные HbS, HbG, НЬС, НЬН и т. д. Общность всех гемоглобинов — в способе укладки их полипептидных цепей вокруг большого плоского кольца гема, идентичного для всех, в центре которого находится атом железа (порфириновое кольцо).
Г ем состоит из атомов углерода, азота и водорода, образующих плоское кольцо, называемое порфирином (рис. 3.17). В центре кольца находится атом Fe, связанный с атомами кольца четырьмя координационными связями (из шести возможных). К гему примыкают два остатка гистидина (His). Имидозольная группа гистидина (F-8) связана координационной связью с атомом Fe через пятую координационную связь. Шестая связь служит для соединения с молекулой О2.
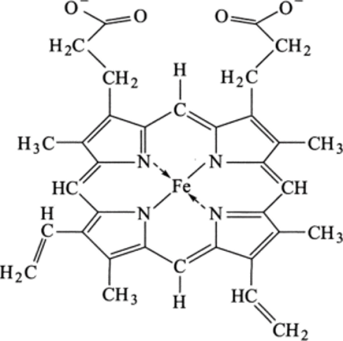
Рис. 3.17. Химическая структура гема.
Миоглобин — мышечный белок, переносящий кислород в мышечных клетках. Он состоит из одной полипептидной цепи, содержит только а-спирали, соединенные петлями, и имеет один гем. Аминокислотная последовательность миоглобина отличается от последовательностей a-цепей гемоглобина. Однако третичная структура a-цепей гемоглобина и миоглобина идентична. Общий способ свертывания а-спиралей глобулярных белков называется глобиновым типом сворачивания.