Передвижение почвенного раствора в растениях под влиянием испарения воды

Исследование ближайших причин передвижения пасоки н почвенного раствора по растению. Основываясь на предыдущих опытах, большинство исследователей (Гофмейстер, Унгер и Сакс) приписывают всасывание пасоки и поднятие почвенного раствора по растению деятельности корней; стебель же они приравнивают наставленной на корень трубке, которая лишь проводит жидкость вверх по древесине для покрытия потери… Читать ещё >
Передвижение почвенного раствора в растениях под влиянием испарения воды (реферат, курсовая, диплом, контрольная)
Мы видели, что воздушные части растений беспрерывно теряют воду через испарение и что убыль воды восполняется при посредстве корней из почвы. Испарение становится особенно сильпым в теплое время года, после распускания листьев, и вместе с ним значительно усиливается всасывание водного почвенного раствора корнями. Из корней жидкость разливается по всему растению и доходит до последних его разветвлений; этот ток жидкости, поднимающийся вверх по растению, обозначают названием восходящего сока этим же именем называют жидкость, переполняющую растепие весною и вытекающую из надреза. Жидкость, всасываемая корнями вследствие испарения воды листьями, отличается от пасоки по своему составу; она не содержит органических соединений и представляет лишь весьма слабый водный раствор неорганических солей.
Исследование пути передвижения почвенного раствора. Выше было указано, что кольцеобразная вырезка коры не задерживает развития частей растения выше вырезки; все количество воды и сырой пищи, необходимое для восполнения испаряемой воды и для питания этих частей, беспрепятственно доставляется им по древесине. С другой стороны, вырезывание древесины при возможно тщательном сохранении коры влечет за собою, напротив того, как показали опыты Дюгамеля и Тревирапуса , 89, быстрое отмирание части дерева выше вырезки.
При нормальных условиях восходящий ток жидкости поднимается в растении по древесине параллельно оси ствола или ветви. Гартиг сделал это очевидным, просверлив через ствол дерева два взаимно перпендикулярных, перекрещивающихся сквозных отверстия; затем три отверстия этой крестообразной вырезки были наглухо замкнуты пробками, четвертое же было соединено с сосудом, содержавшим окрашенную жидкость. Жидкость быстро всасывалась растением; несколько времени спустя дерево было срублено и распилено поперек, но всей длине на цилиндрические отрезки. При этом обнаружилось, что вся всосанная жидкость заключалась в части дерева выше вырезки и окрашивала древесину; окрашенные ею места соответствовали крестообразной вырезке древесины, и везде, куда проникла окрашенная жидкость, виднелся на поперечном разрезе красный крест; часть ствола ниже вырезки осталась попрежнему совершенно бесцветною.
Г еле сом, однако, уже было указано, что обусловленный испарением ток воды (почвенного раствора) может пробегать древесину в разных других направлениях, если воспрепятствовать перемещению жидкости по направлению, параллельному оси ствола или ветви. Гелес[1][2] произвел на отрезанной ветви дуба две глубокие, диаметрально противоположные вырезки до сердцевины на расстоянии 4 дюймов одну от другой; ветвь, погруженная нижним концом в воду, оставалась свежею, несмотря на значительное испарение воды. Подобный же результат получился при вырезывании из ствола четырех накрест поставленных одна под другую вырезок, доходящих до самой сердцевины.
Сюда же относится опыт Гелеса с тремя рядом стоящими яблонями. Среднее дерево было привито концами некоторых ветвей к ветвям остальных двух деревьев. После окончательного сращения привитых ветвей со смежными деревьями корень среднего осторожно вынут был из земли и оставлен на воздухе. С этого времени среднее дерево могло получать необходимую влагу и пищу только при посредстве привитых ветвей, вытягивая ее из сращенных с ним деревьев. Тем не менее оно не только оставалось долгое время живым, но и разрасталось, несмотря на то что питательная жидкость протекала и двигалась в древесине по направлению, диаметрально противоположному нормальному.
В последнее время сходный результат был получен Урихом , 9‘ над буком, который оказался сросшимся посредством одной из ветвей со смежным буком; ветвь отходила от ствола на высоте 11 м и была сращена со смежным деревом на расстоянии 60 см от основания; она имела п этом месте от 8 до 9 см в диаметре.
В 1857 г. ствол этого бука был отделен от корня и оставлен висящим на ветви соседнего дерева. Он тем не менее остался живым и в следующие 20 лет разросся значительно в толщину, увеличив свой диаметр на 5 см; при этом он весил приблизительно 460 кг. В это же время бук, на котором висел последний, тоже разросся в толщину на целых 9 см, несмотря на то что значительная часть жидкости, всасываемой его корнями, потреблялась буком, висевшим на его ветви.
Определить элементы древесины, преимущественно проводящие вверх, но растению почвенный раствор, до сих пор не удалось вследствие непригодности употребленных для этой цели приемов. Старались разрешить этот вопрос двумя способами: 1) в растение вводили последовательно две жидкости, дающие между собою осадок, и из мест отложения осадка заключали о путях передвижения жидкости по растению или же 2) заставляли растение всасывать непосредственно окрашенные жидкости. В первом случае поливали почву последовательно раствором железистосинеродистого калия и железного купороса или, оставив растение некоторое время в почве, политой раствором железисто-синеродистого калия, вырезывали из растения пластинки, которые обрабатывали раствором железного купороса. Прием этот оказался, однако, неудобным вследствие того, что железисто-синеродистый калий чрезвычайно вреден для растения. Верхушки корней оказывались отмершими, вследствие чего жидкости была предоставлена возможность проникать в растение через обнаженные сосуды, т. е. иным путем, чем в растениях нормальных.
Второй прием — непосредственное всасывание растением окрашенных жидкостей — также оказался непригодным. Проникание внутрь пигмента и окраска тканей растений получались лишь в опытах с отрезанными ветвями или растениями, у которых случайно были повреждены корни; растения с корнями цельными принимали в себя, как уже показал Гар— тиг*92, лишь воду без пигмента, так что никакой окраски тканей не происходило. Этот факт был подтвержден впоследствии Балионом193 и Бове 194. Ввиду невозможности получения вышеописанными путями желаемого результата я не считаю нужным останавливаться на произведенных в этом направлении работах.
Определение скорости передвижения почвенного раствора. Неоднократные попытки определить скорость передвижения поднимающейся по растению жидкости также еще не увенчались успехом. Первые опыты с этой целью произведены были уже Гелесом он вычислил скорость поднятия воды по стеблю подсолнечника из сопоставления количества испа-[3][4][5][6]
рясмой воды и площади поперечного разреза стебля. Подсолнечник испарял в продолжение 12 часов 34 кубических дюйма воды; поперечный разрез ствола занимал площадь в 1 квадратный дюйм; если предположить, что стебель полый, то в 12 часов через него пришел бы столб воды в 34 дюйма высоты. По приблизительному расчету, Г елее принял, что '/з разреза занята твердою массою и что только */а оставались свободными для движения воды, откуда он вывел, что в 12 часов должен был пройти через разрез стебля столб воды в 45*/* дюймов; при перечислении на секунды и миллиметры это число дает скорость, равную 0,03 мм в секунду. Сакс195 справедливо замечает, что величина, найденная Гелесом, гораздо меньше действительной; по вычислению Сакса, нужно принять в данном случае скорость приблизительно в 3 раза большую.
Своеобразный способ для определения скорости восхождения воды по растению был предложен Пфицером[7][8][9][10]. Он оставлял растение без поливки почвы до тех пор, пока оно не начинало завядать. Затем почва смачивалась водою и наблюдалось время, когда листья начинали вновь приподниматься. Выводы из подобных опытов были признаны, однако, самим Пфицером сомнительными ввиду того, что поднятие листьев могло быть вызвано водою, находившеюся в растении до поливки почвы, и передвинутой только несколько вверх по растению напором жидкости, вошедшей в корни после смачивания почвы водою.
Третий способ определения скорости поднятия воды по растению принадлежит Мек-Небу 197 и состоит в том, что вводят в растение раствор соли лития, легко узнаваемой в минимальных количествах, но окраске пламени при сожигании вырезанных из растения на различной высоте пластинок. К сожалению, полученные им результаты оказались неверными по двум следующим причинам; во-первых, он производил все опыты не над цельными растениями, а над отрезанными, неукоренившимися ветвями; во-вторых, найденная скорость поднятия в час времени представляет величины вычисленные, а не полученные непосредственно из опыта. Пфицер, проверявший опыты Мек-Неба, впал в те же ошибки. Подобно Мек-Небу, он погружал отрезок ветви (иногда срезываемой в растворе соли лития) нижним концом в воду и затем, но прошествии одной или нескольких минут вырезывал из ветви поперечные пластинки ткани и расследовал на содержание лития. Зная время, прошедшее от погружения конца ветви в раствор соли лития до вырезывания пластинок, можно было определить скорость всасывания раствора. Полученные величины Мек-Неб и Пфицер перечисляли на час времени. Этим приемом Мек-Неб определил наибольшую скорость поднятия в ветви Prunus Lauro-Cerasus равною 1,02 м в час. По Пфицеру, скорость поднятия раствора лития по растению гораздо большая; в некоторых случаях она достигала, по его определению, 33 м в час. Получение таких чрезмерпых цифр становится понятным, если только обратить внимание на открытое Гёнелем разрежение газов в замкнутой отовсюду сети полостей сосудов; неправильность перечисления высот поднятия раствора лития в первые минуты опыта на час времени не требует дальнейших разъяснений.
Сознавая ошибки в исследованиях Мек-Неба и Пфицера, Сакс[11] усовершенствовал этот прием, поливая раствором соли лития почву, в которой выращивались растения. Но и Саксу не удалось окончательно выяснить этого вопроса; открытие лития в золе растения с желаемою точностью удавалось лишь при поливке почвы раствором соли лития в 1—3%, т. е. при концентрации, непригодной для нормального питания растения; при употреблении же раствора более слабого реакция на литий получалась неопределенная. Этим обстоятельством, вероятно, объясняются значительные разницы в скорости поднятия жидкости, которая в опытах Сакса колебалась в пределах между 18 и 206 см в час. Во всяком случае, эти цифры гораздо больше подходят к искомым, чем числовые данные Мек-Неба и Пфицера.
Исследование ближайших причин передвижения пасоки н почвенного раствора по растению. Основываясь на предыдущих опытах, большинство исследователей (Гофмейстер, Унгер и Сакс) приписывают всасывание пасоки и поднятие почвенного раствора по растению деятельности корней; стебель же они приравнивают наставленной на корень трубке, которая лишь проводит жидкость вверх по древесине для покрытия потери воды через испарение. Питра, напротив того, не считая этого взгляда доказанным, поставил себе главною задачею расследовать, не может ли проявляться всасывание жидкости и выделение ее из верхнего конца в отрезках воздушных частей растения, например в кустах стебля, ветвей или в отдельных листьях. Первые его опыты[12][13] состояли в том, что он погружал срезанные ветви с листьями различных древесных пород в воду, оставляя поверх воды только срезанный конец ветви, и прикреплял к нему посредством каучука вертикальную стеклянную трубочку. По прошествии более или менее продолжительного времени появлялся над верхним концом отрезка столб воды, достигавший во многих опытах весьма значительной длины 20°.
Ветвью липы был выделен столбик воды в 70 мм длины «сосны «* * «234,5 * «.
" дуба «» * * 83 * «.
" вишни «» «» 217,5 ««.
* груши * «» «54 «>
" ивы (Saliх «» «» 41 ««.
alba).
Ветви вбирали воду листьями и проводили ее в осевую часть ветви, из верхнего среза которой она выделялась.
Примечание. По мнению Питра, эти опыты доказывают, между прочим, всасывание листьями воды в капельно-жидком виде; во многих опытах, однако, листья, по его показанию, оказывались сгнившими, так что остается нерешенным, насколько всасывание воды в данном случае являлось нормальной функцией листьев.
Подобное же накопление сока в надставленной трубочке обнаружили отрезки различных растений, покрытые соцветиями или несозревшими плодами, также (у некоторых растений) отрезки ветвей, с которых листья были предварительно удалены. Отдельные листья (Acacia heterophylla, Sorbus Aucuparia) производили лишь весьма слабое выделение воды; Питра поэтому приписывает преобладающую роль в выделении жидкости осевым частям отрезков ветвей. Слабое всасывание отрезков березы и сосны после срезывания листьев Питра объясняет особенностями их организации, так как у других древесных пород (рябины, вишни) удаление листьев не задерживало выделения воды. Так, например, в опыте 15 лишенная листьев ветвь Prunus Cerasus выделила столб воды в 142 мм[14].
Опыты с отрезками многолетних ветвей, лишенных почек и листьев, дали сходный результат. Ветвь погружалась в воду верхушкою; при этом нижний конец ее предварительно затягивался каучуком; чтобы облегчить воде доступ в древесину, Питра снимал на известном протяжении кору, а с остальной поверхности ветви удалял пробковую ткань.
Для определения силы, с которой выделялась жидкость из отрезка, наставная трубочка с водою заменялась манометром со ртутью.
Давление, под которым вытекала жидкость, выразилась в следующих цифрах[15]:
Отрезок Betula alba поднял ртуть на 75 мм.
* Sorbus Aucuparia «» 49 *.
" Prunus Cerasus «» 116 «.
" Pinus sylvestris «» 114 *.
* Populus pyramidalis «» 30 «.
Просматривая список растений, отрезки которых выделяли воду, нельзя не заметить, что между ними находятся растения, которые при нормальных условиях никогда не обнаруживают плача, например хвойные и тополь. Питра приписывает выделение жидкости отрезками ствола и ветвей осмотическим процессам и считает доказанным активное участие их в поднятии жидкости по растению.
Подтверждая все факты, приведенные Питра, Бем[16] оспаривает его выводы и заканчивает критическую заметку заявлением, что в стеблевых частях растений нет (осмотических) сил, выдавливающих пасоку, подобно тканям корней. Выделение жидкости отрезками воздушных частей растений Бем объясняет совершенно иными процессами; он предполагает, что погруженный в воду отрезок всасывает в первое время большое количество воды; затем загнивает, внутри его обнаруживается масляное брожение, сопровождаемое выделение газов, преимущественно углекислоты и водорода. Бем полагает, что выделение отрезком воды и поднятие в трубочке жидкости обусловливается газами, которые развиваются в отрезке при гниении.
С мнением Бема, однако, трудно согласиться; между опытами Питра имеются и такие, где выделение жидкости наступало сравнительно скоро и образование газов масляного брожения является весьма сомнительным. Я со своей стороны склоняюсь на сторону Питра и надеюсь, что при дальнейшей разработке вопроса выводы его подтвердятся.
Примечание. К воззрениям, сходным с высказанными Питра, пришел К. Краус[17][18][19][20]; он также принимает в надземных частях растений силу, которая гонит жидкость кверху и содействует ее поднятию. Вытекание сока Краус, как и Питра, наблюдал на отрезках растений; из описания его, однако, нельзя с достоверностью заключить о характере описанного им явления. Он наблюдал вытекание жидкости только по каплям; кроме того, она преимущественно выделялась из камбия и ситовидных элементов и никогда не вытекала из сосудов; на пасоку не походила она и по консистенции. Ввиду этих соображений выводы Крауса являются весьма сомнительпыми, тем более что во многих случаях выделение капли жидкости наблюдалось им в то время, когда отрезок, видимо, начал приходить в гниение.
Попытки разгадать и объяснить поднятие жидкости по растению не привели еще к определенным результатам. Некоторые физиологи (Гофмейстер) 205 старались объяснить восхождение воды по растению эндосмотическими процессами, другие (Жамен) 208 — волосностью, третьи (Бем) 207 — совокупным действием упругости оболочек клеток, давления атмосферы и испарения, четвертые (Сакс) приписывают поднятие воды по древесине скважности 4* последней. Мнения также расходятся относительно участия тканей ствола в поднятии жидкости; большинство физиологов полагают, как мы видели, что ткани ствола не принимают активного участия в поднятии жидкости и служат только проводниками ее в испаряющие органы, между тем как некоторые (Питра) приписывают стволу и ветвям такое же активное участие в этом процессе, как и корням.
С целью подкрепить теоретические соображения фактами были устроены Гофмейстером, Жаменом и Бемом приборы, долженствовавшие функционировать по отношению к поднятию жидкости подобно растениям, сообразно со взглядом автора. Не придавая решающего значения ни одной из предлагаемых теорий поднятия жидкости по растению, я ограничиваюсь вышеприведенным указанием статей, в которых они изложены, и перехожу к фактической разработке этого вопроса.
Выше была выяснена активная роль корней в поднятии жидкости и показано, что деятельность их продолжается за весь период вегетации, до поздней осени, с тою только разницею, что начиная с лета количество жидкости, вводимое активною деятельностью корней, постепенно убывает, между тем как сила, с которою корни выталкивают воду в надземные части, остается весьма значительною вплоть до прекращения вегетации.
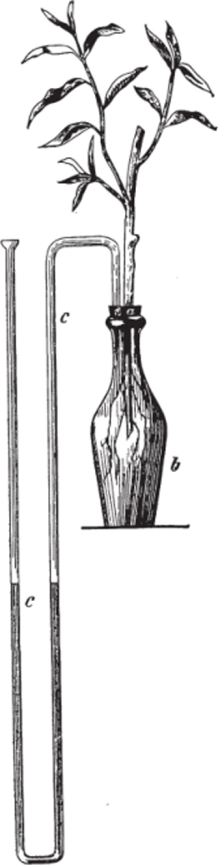
Рис. 91.
Относительно участия надземных, частей в поднятии жидкости имеются следующие указания: Г елее[21] уже заметил, что отрезанные ветви всасывают воду со значительною силою; для измерения ее он прикреплял ветвь к стеклянной трубке и, наполнив трубку водою, опускал ее нижним концом в ртуть. По мере всасывания воды ветвью ртуть в трубке поднималась на высоту нескольких дюймов; в опытах с ветвями яблони и виноградной лозы столб ртути достигал длины от 4. до 11 дюймов; этим не измерялась еще вполне сила всасывания воды, так как из нижнего конца ветви выделялись во множестве пузырьки газа, которые, скопляясь в верхней части трубки, уединяли ветвь от воды и представляли преграду дальнейшему ее всасыванию.
Подобное же сильное всасывание воды наблюдал Гелесу прикрепив трубку с водою к дереву в месте, с которой была срезана ветвь; ртуть иногда уже через б мин поднималась на 8 дюймов.
Еще более сильное всасывание воды получил Гартиг[22] в опытах над Carpinus betulus; в боковую цилиндрическую вырезку ствола он вставлял, по возможности плотно, тубулированную реторту, наполненную водой; в отверстии реторты была прикреплена трубка, налитая доверху водою. Опрокинув реторту трубкою вниз, Гартиг вводил горлышко реторты в ствол, а открытый (нижний) конец трубки погружал в сосуд с ртутью. Вода всасывалась с такою силою, что поднимала ртуть на 17 дюймов, т. е. высоту соответствующую 2/3 атмосферного давления; но и в этом случае дальнейшему поднятию ртути препятствовал воздух, перешедший из ствола в реторту.
Для устранения этого препятствия при наблюдении силы всасывания воды растениями Бем2,0 видоизменил опыт следующим образом (рис. 91): укоренившаяся ветвь вставлялась в плотно прилаженную пробку (а) сосуда (Ь) так, чтобы нижний конец ее и корни приходились на значительной глубине жидкости;
выделявшиеся из ветви пузырьки воздуха не могли, следовательно, препятствовать всасыванию; сила всасывания измерялась прилаженным манометром ©. Покрытая листьями ветвь поднимала иногда в солнечный день ртуть до 60 см.
Не выясняя причины всасывания воды растепием, эти опыты, однако, несомненно свидетельствуют о сильном всасывапии воды надземными частями растения, обусловленном главным образом испарением. Участие тканей ствола и ветвей ограничивается, по всему вероятию, только легкою проницаемостью древесины для воды, т. е. отсутствием препятствий для перемещения ее в верхние, испаряющие части растения. Относительно проводимости воды древесипою имеются интересные опыты Саксаги он показал при этом, что способность ткани проводить воду не находится в связи с разбуханием ее в воде; ткани, наиболее разбухающие в воде, оказались весьма дурно проводящими воду. Если высушенную на воздухе Laminaria, которая чрезвычайно сильно разбухает в воде, опустить в воду нижним концом, то в погруженной части она сильно разбухает, на всем же остальном протяжении вне воды водоросль остается по-прежнему совершенно сухою; свежая Laminaria также не вбирает в себя воды в части, оставленной на воздухе, и засыхает на всем протяжении поверх воды.
Сильно разбухающие вещества, как, например, животный клей, свернувшийся белок, обнаруживают то же самое. Крахмал, жадно впитывающий воду в сухом состоянии, переставал пропускать ее по превращении в клейстер. Сакс доказал это, погружая в горячую воду стеклянную трубку, плотно набитую крахмалом и завязанную снизу куском холста, чтобы воспрепятствовать высыпанию крахмала. Горячая вода немедленно превращала нижний слой крахмала в клейстер, который препятствовал дальнейшему прониканию воды в трубку, так что крахмал в ней оставался сухим, несмотря на то что нижний конец трубки находился па глубине 30—50 см под поверхностью воды[23]‘[23].
Древесина оказалась, по Саксу, отличною от других тканей: 1) по сравнительно малому количеству воды, потребному для пресыщения, и 2) способностью пропускать сквозь себя воду с большою скоростью.
Сакс принимает, что вода фильтруется по стенкам клеток древесины, оставляя в стороне их содержимое: кусок древесины Abies pectinata в 100 см и более длины, вырезанный зимою, когда ткань ее очень богата водою, столь быстро пропускал воду, что капля воды, перенесенная на верхний конец отрезка, всасывалась и в то же время немедленно вызывала появление капли воды на нижнем конце отрезка. При переворачивании отрезка верхним концом вниз капля вновь исчезала на верхнем и образовалась на нижнем конце. Для обнаружения скорости фильтрования воды через древесину Сакс прикреплял к короткому концу трубки, изогнутой в виде буквы U, отрезок древесины; в трубку наливалась затем вода, так чтобы в длинном колене она стояла выше места прикрепления отрезка в коротком колене; под давлением этого столба воды жидкость фильтровалась сквозь отрезок и собиралась в надставленной над отрезком трубке.
Через отрезок Taxus baccata под давлением столба воды в 70 мм проходил в 2 мин столб воды в 50 мм высоты, что составляет в 1 час времени столб воды в 1,5 м высоты; отрезок Abies pectinata пропускал воду со скоростью 690 мм в час при давлении столба воды в 68 мм длины. Вода фильтровалась преимущественно через осенние слои древесины. Приведенные числа выражают быстроту фильтрования воды в древесине по оси ствола или ветви; она оказалась гораздо меньшею в горизонтальном направлении; кусок древесины Abies pectinata в 48,5 мм длины, вырезанный поперек ствола по радиальному направлению, не пропустил ни одной капли воды в первые часы опыта; несмотря на давление столба воды в 160 см, за 24 часа прошло сквозь него 2,3 см3 жидкости.
Трудность, с которой фильтровалась вода по радиальному направлению, объясняется частью отсутствием окаймленных продушин ** на тангептальных стенках, частью тем, что вода, двигаясь по этому направлению, должна пройти через много слоев осенней древесины, трудно пропускающей воду.
Опыты Сакса особенно интересны, так как относятся до растений, совершенно лишенных сосудов; трахеиды древесины хвойных представляют, по согласному свидетельству всех исследователей, клетки с оболочкою отовсюду замкнутою, без отверстий. Проводимость древесины оказалась, следовательно, вне связи с сосудами, которые, за исключением сердцевинной трубки, совершенно отсутствуют в хвойных 6*.
Из всего вышесказанного выяснились следующие результаты касательно передвижения по растениям газообразных, жидких и твердых тел. Все выводы относятся только к высшим цветковым растениям.
- 1. В растении происходят беспрерывно обмен и перемещение газообразных, жидких и твердых тел.
- 2. Газообразные тела заключены в воздушных вместилищах, которые представляют две системы ветвистых полостей, разобщенных между собою; одну из них составляют сосуды древесины, другую — межклетники коры и сердцевины, сообщающиеся между собою посредством сердцевинных лучей.
- 3. Вторая из этих двух систем полостей, т. е. сеть межклетников коры (и сердцевины) у водных растений, вследствие отсутствия устьиц остается уединенною от окружающей атмосферы; у сухопутных, напротив того, сообщается в надземных частях растений с воздухом при посредстве устьиц, которые местами столь многочисленны, что, несмотря на микроскопические размеры, поддерживают свободный обмен газов между растением и атмосферой.
Этою анатомическою особенностью объясняется различие в составе газов этих полостей.
У водных растений, где обе системы полостей не только разобщены между собою, но и уединены от внешней среды, каждая из них может по временам заключать смесь газов, различную и не сходную со смесью газов вне растения. Подобные примеры приведены мною выше.
В растениях сухопутных, напротив того, только в системе полостей сосудов древесины наблюдается смесь газов, отличная от воздуха как по давлению, под которым газы в ней находятся, так и по составу; система же межклетников коры достаточно быстро вентилируется при посредстве устьиц, и заключенные в ней газы не разнятся значительно от атмосферного воздуха. Помимо этих полостей, газы перемещаются по тканям растений вследствие диффузии.
- 4. Не менее разнообразны пути передвижения по растению жидких и твердых тел. Они управляются, с одной стороны, причинами внутренними, с другой — внешними. Под влиянием внутренних причин происходят, например, перемещения пластического материала из мест его образования в места потребления или временного отложения и из последних опять в молодые нарастающие органы. Этого рода передвижения воды и растворенных в ней тел свойственны всем растениям. К ним присоединяется у сухопутных растений (и у водных, которые верхними частями выставляются из воды) еще весьма значительный ток почвенного раствора, пробегающий по растению снизу вверх и управляемый исключительно испарением. Из разнообразнейших направлений, по которым происходит перемещение жидких и твердых тел, но растению, исследованы более обстоятельно четыре пути: а) перемещение запасного материала семени при прорастании из белка и семядолей в нарастающие части зародыша; б) передвижение вниз по коре пластического сока, вырабатываемого листьями; в) передвижение в древесных породах зимнего запасного материала из ветвей, ствола и корня в весенние нарастающие побеги, а также и тесно связанное с ним у некоторых растений явление плача и г) пробегающий снизу вверх по растению ток почвенного раствора, обусловленный главным образом испарением.
- 5. Точных указаний на ткани, служащие проводниками главнейших составный частей пластического сока, до сих пор получить не удалось. Неизвестно даже с достоверностью, в виде каких соединений передвигается пластический материал. Но отношению к азотистым соединениям доказано, что они передвигаются не только в виде белковых тел, но и в виде амидов, между которыми чаще всего встречается аспарагин. Подобное предположение было высказано в последнее время также касательно углеводов ввиду трудной проницаемости гиалоплазмы по отношению к сахару, а также и вследствие полпого отсутствия сахара и декстрина при прорастании некоторых семян, содержащих крахмал.
Поэтому выводы Сакса о передвижении углеводов по паренхиме и азотистых тел в виде белковых соединений по ситовидным элементам и камбиформу, а равно и показания Бриози и Ганштейпа касательно перемещения пластического сока в полном составе по ситовидным элементам потеряли в настоящее время прежнее значение.
6. Всасывание растением почвенного раствора и передвижение его по растению производятся частью деятельностью корней, главным же образом управляются испарением воды растением.
- [1] Treviranus. Physiolog. d. Gewachse, В. 1, p. 286 (1835).
- [2] Hales. L. c., p. 115.
- [3] Urich. Forstwissensch. Centralbl. v. Baur, p. 468. (Just. Jahresb. 1879, p. 292).
- [4] Hartig. Bot. ZeiL 1853, 309.
- [5] Haillon. C. R. 80, 426 (1875).
- [6] Cauvet. C. R. 81, 52 (1875).
- [7] Sachs. Experimental physiol., р. 234 (1865).
- [8] Pfitzer. Verhandl. d. naturhisL Vereins z. Heidelberg.
- [9] Mac-Nab. Transactions of the botanical soc. of Edinbourgh, Vol. 11; 45 (1871). Его
- [10] же: Transactions of Irisch Academy Dublin, 1874, p. 343.
- [11], 9 В Sachs. Arbeit., d. bot. Instit. z. Wurzburg.
- [12] Pitra. Pringsh, Jahrbuch. II, 437 (1878).
- [13] Pitra. L. c., p. 452 u сл.
- [14] L. с., р. 465.
- [15] L. с., р. 505 и сл.
- [16] Boehm. Bot. Zeit. 1880, р. 33.
- [17] Carl Kraus. Flora, 1881; N 2, 4, 5 и 6.
- [18] Hofmeister. Flora, 1862, 106.
- [19] 209 Jamin. Lemons professees.
- [20] Boehm. Sitzungesb. Wien. Ak. 48 (Separatabdruck) 1863 и 1864. Его же: Land.Vers. 20; 363 (1877).
- [21] 'ов Hales. L. с., р. 87. t09 Hartig. Bot. Zeit, 1861, p. 18.
- [22] 2,0 Boehm. Sitzungsb. Wien. Ak. 48, (Separatabdr. p. 17) (1863).
- [23] 21 ‘ Sachs. Arbeit d. bot. Instit z. Wurzburg, p. 201.4,2 Sachs. L. c., p. 314.
- [24] 21 ‘ Sachs. Arbeit d. bot. Instit z. Wurzburg, p. 201.4,2 Sachs. L. c., p. 314.