Регуляция синтеза белка у эукариот

Регуляторными элементами являются белки инициаторного комплекса, описанные в гл. 28. В дополнение можно отметить особые нуклеотидные последовательности, способствующие интенсификации транскрипции, так называемые энхансеры. Характерная особенность этих структур заключается в том, что они влияют на скорость транскрипции независимо от локализации в опероне. Белки, взаимодействующие с энхансерами… Читать ещё >
Регуляция синтеза белка у эукариот (реферат, курсовая, диплом, контрольная)
Это более сложный процесс, так как транскрипция и трансляция происходят в разных компартментах и обеспечиваются большим количеством соответствующих структур.
На уровне транскрипции регуляторные механизмы у прокариот и эукариот имеют ряд обших черт. Рассмотрим некоторые отличительные особенности. Для клеток эукариот характерна амплификация генов и их перестройка. Оба механизма обеспечивают резкое увеличение копий тех или иных белков, необходимых для реализации клеточного метаболизма.
Известно, что в клетках эукариот ДНК, соединенная с белками (гистонами), упакована в нуклеосомы (гл. 14). В этом состоянии транскрипция невозможна, и для экспрессии генов необходимо деблокирование транскриптона. Следовательно, образование и разрушение нуклеосом является важным фактором регуляции эукариотических генов. Каким же образом происходит деблокирование транскриптона?
Фосфорилирование гистонов. В результате действия белковых гормонов происходит опосредованное фосфорилирование ядерных белков — гистонов и разрушение нуклеосом. Матрица при этом становится доступной для основных факторов инициации транскрипции, и начинается синтез РНК. При прекрашении действия гормонов нуклеосомы восстанавливаются.
Ацетилирование и деацетилирование гистонов. Это важный фактор регуляции генной активности. Оказалось, что фермент гистон-ацетилаза ассоциирована с фактором ТАФ (гл. 28). Ацетилирование проходит по терминальному остатку лизина в полипептидной цепи гистона. В результате ацетилирования положительный заряд белка уменьшается и сродство гистона к отрицательно заряженной ДНК снижается. Это может привести к разрушению нуклеосом и деблокированию транскриптона. Деацетилирование гистонов приводит к противоположному эффекту. Специфические ацетилаза и деацетилаза ассоциированы с белками инициации транскрипции.
Регуляторными элементами являются белки инициаторного комплекса, описанные в гл. 28. В дополнение можно отметить особые нуклеотидные последовательности, способствующие интенсификации транскрипции, так называемые энхансеры. Характерная особенность этих структур заключается в том, что они влияют на скорость транскрипции независимо от локализации в опероне. Белки, взаимодействующие с энхансерами, называются энхансерными элементами, расположенными на расстоянии 1000—2000 пар оснований от региона промотора. Эти белковые факторы способны воздействовать на инициацию транскрипции благодаря образованию ДНК-петли, что приводит к пространственному сближению энхансерных элементов и, например, белков ТАТА (рис. 29.7).
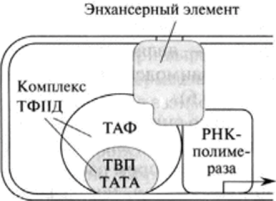
Рис. 29.7. Взаимодействие энхансериого элемента с белками инициаторного комплекса.
Весьма существенным фактором регуляции транскрипции является процессинг РНК. Образование зрелых мРНК зависит от скоростей кэпирования, образования полиА, а также скорости стшйсинга. Для полицистронных мРНК определенное регуляторное значение имеет альтернативный сплайсинг (гл. 30).
Кроме белков инициаторного комплекса, на скорость транскрипции оказывают существенное влияние ДНК-связывающие белки. Из нескольких семейств наиболее известны белки типа: цинковые пальцы, спираль—виток—спираль и гомеодоменные белки. Специфическое связывание этих белков с ДНК происходит в результате взаимодействия боковых радикалов аминокислотных остатков белка с основаниями ДНК.
Цинковые пальцы представляют собой серию повторяющихся доменов (от двух до девяти), имеющих форму пальца. В центре координации каждого домена находится цинк. В одних случаях цинк соединен с четырьмя остатками цистеина, в других — с двумя цистеинами и двумя гистидинами.
На конечный результат — синтез белка — влияет также скорость транспорта РНК в цитоплазму. В цитоплазме мРНК, взаимодействуя с определенными белками, образует информосому — своеобразное депо, из которого мРНК освобождается по мере надобности для синтеза белка. Скорость освобождения мРНК также является фактором регуляции белкового синтеза.
Скорость синтеза белка напрямую зависит от количества мРНК, которое определяется временем ее «полужизни» или стабильностью in vivo. Таким образом, факторы, влияющие на стабильность мРНК, являются регуляторами экспрессии генов и, как следствие, белкового синтеза. Одной из структур, определяющих стабильность мРНК, является полиА-последовательность на З’ОН-конце. Факторы, регулирующие экспрессию генов и синтез белка, суммированы в табл. 29.2.
Таблица 29.2. Факторы, влияющие на регуляцию транскрипции эукариот.
Число. | Факторы. | Число. | Факторы. |
| Амплификация генов Перегруппировка генов Белки ининиаторного комплекса ДНК-связываюшие белки. |
| Сплайсинг мРНК Стабильность мРНК Транспорт мРНК в цитоплазму. |
Лимитирующей стадией процесса трансляции является ее инициация. Наиболее подробно описан процесс изменения скорости инициации трансляции в результате фосфорилирования фактора инициации IF2. Реакция катализируется ферментом 1Р2-киназой, причем присоединение фосфатной группы инактивирует фактор инициации. Этот феномен был изучен на примере синтеза гемоглобина в ретикулоцитах. Сначала было установлено, что глобин синтезируется только в присутствии гема. Затем была выстроена вся система регуляции синтеза глобина. Оказалось, что активация 1Р2-киназы происходит за счет ее фосфорилирования цАМФ-зависимой протеинкиназой. Взаимодействие этой протеинкиназы с цАМФ и ее активацию блокирует гем, выполняя тем самым негативный контроль синтеза гемоглобина.
Регуляция синтеза белка осуществляется также на стадии процессинга белка. Модификации новосинтезированных полипептидов осуществляются при помощи соответствующих ферментов, активность которых, в свою очередь, находится под генетическим контролем. К этим модификациям относятся метилирование, фосфорилирование, гликозилирование, а также ограниченный протеолиз.