Механизм окислительного фосфорилирования

Нобелевский лауреат П. Митчел предложил хемиосмотическую модель. Электрохимическая энергия рождается из-за разности концентрации протонов и разности заряда вокруг внутренней мембраны митохондрии. Градиент (Дм, «дельта мю») получил название электрохимический градиент или протонный градиент. Он имеет две составляющие — электрическую (ДШ, «дельта пси») и концентрационную (ДрН): Все атомы водорода… Читать ещё >
Механизм окислительного фосфорилирования (реферат, курсовая, диплом, контрольная)
На основании строения и функций компонентов дыхательной цепи предложен механизм окислительного фосфорилирования:
1. Ферменты дыхательной цепи расположены в строго определенной последовательности: каждый последующий белок обладает большим сродством к электронам, чем предыдущий (он более электроположителен, т. е. обладает более положительным окислительно-восстановительным потенциалом). Это обеспечивает однонаправленное движение электронов.
Все атомы водорода, отщепленные дегидрогеназами от субстратов в аэробных условиях, достигают внутренней мембраны митохондрий в составе НАДН или ФАДН2.
НАД — никотинамид аденин динуклеотид водорастворимый переносчик электронов. Окислительное фософорилирование начинается с поступления электронов в дыхательную цепь. Поставщиками электронов для НАД (НАДФ) служат различные дегидрогеназы цитозоля или митохондрий. У субстрата забираются два атома водорода. Один связывается с НАД, другой выходит в окружающую среду.
Реакция выглядит следующим образом:
Восстановленный субстрат + NAD = окисленный субстрат + НАДН + Н*.
Внутренняя мембрана митохондрий непроницаема для НАДН, поэтому электроны передаются с него непосредственно на акцепторы первого комплекса ЦПЭ.
ФАД — флавин аденин динуклеотид. Флавопротеины обычно очень тесно (ковалентно) связаны с флавиновыми нуклеотидами (ФМН, ФАД), которые часто являются коферментами активного центра. Окисленный получает два электрона и образуется ФАДН 2 или ФМНН 2. В цепь они их могут отдавать по одному электрону.
Митохондриальная дыхательная цепь состоит из серии согласованно работающих переносчиков электронов, большинство из которых представлены интегральными белками, простетические группы которых способны получать и отдавать один или два электрона. Есть три типа переносчиков:
- 1. Прямой перенос электронов путём восстановления Fe3+ в Fe2+;
- 2. Перенос в виде атома водорода Н+ + е-;
- 3. Перенос в виде иона водорода: Н-, который несёт два электрона.
Кроме НАД и флавопротеинов переносчиками электронов служат:
· гидрофобная молекула хинона (убихинон, коэнзим Q, коэнзим Q10). Это жирорастворимый бензохинон с длинной боковой изопреноидной цепью (у человека — 10 едениц). Наподобие флавопротеинов служит связующим звеном между двухэлектронными донорами и одноэлектронными акцепторами электронов.
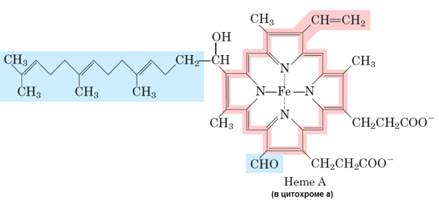
Существует два различных типа железосодержащих белков:
· цитохромы; Митохондрии имеют три вида цитохромов а, в и с. Цитохромы а и в нековалентно связаны с белком, который встроен во внутреннюю мембрану митхондрий, а с — ковалентно связан с цистеином цитохромов. Цитохром С исключительный — растворимый белок цепи, который электростатически связан с внешней поверхностью внутренней мембраны.
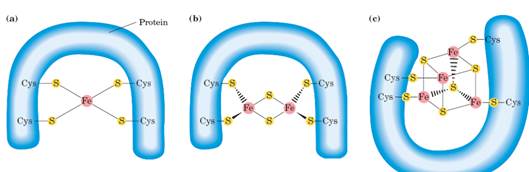
· железосерные белки (Fe-S). Атом железа связан с гемом, а с неорганической серой или серой в составе цистеина белка.
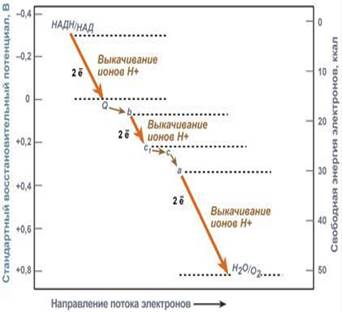
Здесь атомы водорода (от НАДН и ФАДН 2) передают свои электроны в дыхательную ферментативную цепь, по которой электроны движутся (50−200 шт./сек) к своему конечному акцептору — кислороду. В результате образуется вода.
Поступающие в дыхательную цепь электроны имеют высокий энергетический уровень. По мере их продвижения по цепи они теряют энергию.
Энергетические соотношения в дыхательной цепи митохондрий и участки переноса ионов Н+ через мембрану. Часть энергии электронов используется I, III, IV комплексами дыхательных ферментов для перемещения ионов водорода через мембрану в межмембранное пространство. Другая часть рассеивается в виде тепла.
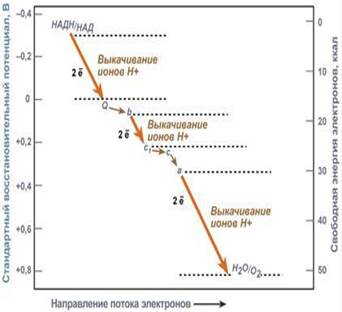
Перенос ионов водорода через мембрану формируется градиент ионов водорода между внутренней и наружной поверхностями внутренней митохондриальной мембраны. Такой градиент обладает потенциальной энергией.
Нобелевский лауреат П. Митчел предложил хемиосмотическую модель. Электрохимическая энергия рождается из-за разности концентрации протонов и разности заряда вокруг внутренней мембраны митохондрии.
Градиент (Дм, «дельта мю») получил название электрохимический градиент или протонный градиент. Он имеет две составляющие — электрическую (ДШ, «дельта пси») и концентрационную (ДрН):
Дм = ДШ + ДрН.
Как завершение всех предыдущих событий и необходимый их результат происходит наработка АТФ: ионы H+ теряют свою энергию, проходя через АТФ-синтазу (Н+-транспортирующая АТФ-аза, КФ 3.6.3.14.). Часть этой энергии тратится на синтез АТФ. Другая часть рассеивается в виде тепла.