Новый аспект парасексуального цикла.
Цитодукция

Более строгий подход к изучению перекомбинации ядер и неядерных элементов разработан для дрожжей-сахаромицетов, у которых при гибридизации также образуется, хотя и кратковременная, стадия гетерокариона. Р. Райт и Дж. Ледерберг (1957) использовали это явление для изучения рекомбинации неядерных детерминант признака Pet и ядерных маркеров. Они показали, что первые почки зиготы дрожжей, полученные… Читать ещё >
Новый аспект парасексуального цикла. Цитодукция (реферат, курсовая, диплом, контрольная)
Существование полуавтономных клеточных органелл со своими генетическими детерминантами, а также присутствие в клетке других генетических детерминант, локализованных вне ядра, побуждает обратиться еще к одной стороне парасексуального процесса. В ходе этого процесса, инициируемого слиянием гиф и образованием гетерокарионов, могут объединяться различные по генетической характеристике неядерные элементы. Они по-разному комбинируются друг с другом и с ядрами, что отражается на общей картине вегетативного расщепления. Примеры такой перекомбинации ядер и неядерных элементов получены у мицелиальных грибов Neurospora, Aspergillus, Podospora и др. Однако само естественное состояние многоядерного мицелия у этих объектов создает трудности в изучении процесса такого вегетативного расщепления.
Более строгий подход к изучению перекомбинации ядер и неядерных элементов разработан для дрожжей-сахаромицетов, у которых при гибридизации также образуется, хотя и кратковременная, стадия гетерокариона. Р. Райт и Дж. Ледерберг (1957) использовали это явление для изучения рекомбинации неядерных детерминант признака Pet и ядерных маркеров. Они показали, что первые почки зиготы дрожжей, полученные от скрещивания клеток rho~ х г ho*, иногда могут оказаться гаплоидными и нести ядра того или другого родителя, судя по рецессивным маркерам, в цитоплазме rho*. Это явление образования ядерно-цитоплазматических гибридов И. А. Захаров (1969) назвал цитодукцией.
Цитодукция оказалась удобной для локализации генов в ядре или цитоплазме, особенно в тех случаях, когда они обнаруживают неменделевское наследование в мейозе. Цитодукция обычно происходит с частотой менее 1%, т. е. менее 1% всех образующихся зигот отпочковывают цитодуктанты — гаплоидные клетки со смешанной цитоплазмой. В экспериментах по цитодукции используют клетки одного родителя, маркированные по митохондриям нейтральной (не супрессивной) мутацией rho0. Этот же штамм обычно несет какую-либо рецессивную ядерную мутацию устойчивости, например к канаванину (сап Г) — аналогу аминокислоты аргинина, или другой рецессивный маркер, по которому легко вести селекцию. Эти клетки — реципиенты цитоплазмы. Второй родитель имеет нормальные митохондрии (rho*) и несет ядерный доминантный аллель чувствительности к тому же агенту (CAN Is). Эти клетки — доноры цитоплазмы. Кроме того, ядра обоих гаплоидов маркируют несколькими неселективными рецессивными мутациями, чтобы можно было следить за поведением хромосом при последующем анализе.
Цитодуктанты отбирают на среде с этанолом (или другим неферментируемым источником углерода) и канаванином при скрещивании (рис. 8.9) донора и реципиента цитоплазмы. На этой среде не могут расти ни донор (он чувствителен к канаванину), ни реципиент (rho0) цитоплазмы, ни нормальный гибрид, который тоже чувствителен к канаванину (CAN 1 s/can l'). Вырастают только гаплоидные цитодуктанты, сохраняющие рецессивную устойчивость и приобретающие нормальные митохондрии (rho*).
Для повышения частоты цитодукции используют ядерные мутации, задерживающие кариогамию. Например, ядерная мутация каг 1 повышает частоту цитодукции до 80% и более.
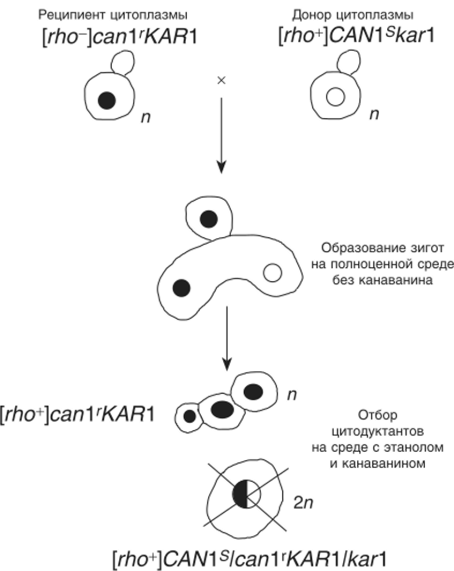
Рис. 8.9. Образование и селекция цитодуктантов при задержке кариогамии у дрожжей.
В опытах Р. Райта и Дж. Ледерберга с помощью цитодукции показана неядерная природа >7ют-детерминантов. В дальнейшем этот метод был использован для установления ядерной локализации криптической (не имеющей своего фенотипического проявления) плазмиды с контурной длиной 2 мкм у дрожжей, а также для установления ядерной локализации сконструированных на ее основе искусственных плазмид с ящерными генами.
Эти плазмиды при цитодукции с митохондриями не передаются и сопровождают ядерные гены того родителя, с которым они введены в скрещивание. При обычном гибридологическом анализе по доминантным плазмидным генам наблюдается резкое преобладание доминантного фенотипа: тетрады 4А: 0<�я или ЗА: 1а. Это объясняется большим числом копий плазмиды в ядре и, соответственно, большим числом копий доминантного аллеля.
Данное обстоятельство следует подчеркнуть особо, т. к. оно показывает, что нарушение типичного хромосомного наследования еще не доказывает неядерную природу исследуемого гена.
Применяя цитодукцию, доказывают цитоплазматическую локализацию двунитевой РНК — вирусоподобного детерминанта убивающей активности, обнаруженного у некоторых штаммов дрожжей-сахаромицетов. Такие дрожжи при совместном выращивании убивают клетки чувствительных штаммов.
Генетический анализ, основанный на цитодукции, широко используется при исследовании дрожжей и может быть распространен на другие объекты.