Математико-статистические методы подбора пар для скрещивания

Общим недостатком методов, оценивающих близость к идеальному сорту средних значений признаков ожидаемого потомства, является отсутствие учета генетического разнообразия популяции потомства. Один вариант скрещивания может давать большую близость средних к идеалу, но иметь низкое генетическое разнообразие популяции потомства по изучаемым признакам, а второй — дальше от идеала по средним… Читать ещё >
Математико-статистические методы подбора пар для скрещивания (реферат, курсовая, диплом, контрольная)
При скрещивании географически отдаленных форм в потомстве ожидается повышенная генетическая изменчивость и, легче обнаружить трансгрессивные рекомбинанты. Географическую отдаленность можно связать с генетической дивергентностью родителей, т. е. с расхождением линий или популяций, происходящих от общего предка, в результате отбора или изоляции. В процессе эволюции в условиях географической отдаленности в местных популяциях закрепляются и накапливаются гены с наибольшей адаптивной ценностью к специфическим условиям среды. У отдельных родителей хозяйственные признаки контролируются различными наборами аллелей, случайные рекомбинации которых вызывают трансгрессивное расщепление.
Поэтому целесообразнее используют термин генетической отдаленности или дивергентности — несходства аллельного состава родительских форм. Основная проблема при оценке степени дивергентности по генам, определяющим различия форм по хозяйственно ценным признакам, — недостаток сведений о наследовании этих признаков, т. е. о связи фенотипических выражений признаков с генотипом. Поэтому часто пытаются оценить общее несходство аллельного состава родительских форм в надежде, что при этом будет учтено различие и по генам хозяйственно — ценных признаков.
Генетическую дивергенцию можно выразить через специальные пара — метры с помощью методов многомерного статистического анализа комплекса количественных признаков.
Естественной мерой отдаленности потенциальных родителей служит евклидово расстояние. Евклидово расстояние между двумя генотипами определяется по теореме Пифагора (2):
(2).
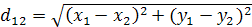
Где xlk, x2k-среднее значение k-го признака у1-го и 2-го сортов.
Для оценки общей генетической дивергентности по всему геному в показатель следует включать максимальное число количественных признаков. Если у признаков различные размерности, то евклидово расстояние может оказаться неверным. Данный метод позволяет выявить группы генетически тесно связанных признаков и отобрать от каких групп по одному признаку для использования в евклидовом расстоянии.
Близость к идеальным значениям по комплексу признаков Для прогноза близости потомства от скрещивания выбранных родителей к заданному идеалу Дж. Графиус разработал векторный метод. Под идеалом понимается модель сорта — воображаемый сорт, имеющий заданные величины признаков, оптимальные для конкретных условий среды.
Д. Педерсон упростил этот метод и предложил подобрать родительские формы, дополнительно задавая селекционным признакам различные веса — допустимые отклонения от идеальных значений. Суть метода Педерсона заключается в следующем. У каждого из т потенциальных родительских сортов учтено N количественных признаков. Номер признака обозначен у. Каждому признаку селекционер — эксперт задает идеальное значение Ij, а также отклонение от идеала Dj. Цель скрещивания — получение гибридной популяции со средним значением каждого признака в интервале Ij± Dj.
Основная проблема прогнозирования — сложная генетическая связь величин признаков родителей и популяционных средних в потомстве их скрещиваний. Поэтому в методе Педерсона предлагается, что среднее значение каждого признака в позднем поколении скрещивания равно средневзвешенным значениям этого признака у родителей. Таким образом, популяционная средняя по j-му признаку (j) потомства равна (3):
(3).
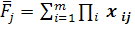
где хij — значение j-го признака у i-го родителя; Пi — доля i-го родителя в геноме популяции потомства. Обычно селекционер располагает большим набором сортов и форм, изучаемых по хозяйственным признакам в конкурсном и предварительном сортоиспытании, производстве или в коллекционном питомнике. Модификация метода оценки близости к идеалу, предложенную С. П. Мартыновым. Мерой отклонения ожидаемого потомства от модели идеального сорта в модификации является уравнение:
(4).
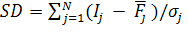
где Ij, j значение jго признака у идеала и ожидаемого потомства; аjвесовoй коэффициент j-го признака; - стандартное отклонение j-го признака в наборе сортов.
В этом случае вектор идеала представляет собой совокупность оптимальных (наилучших) значений селекционных признаков, полученных при испытании сортов в различных условиях.
В методе С. П. Мартынова признаки могут быть разработаны на несколько групп с одинаковыми весами для признаков в одной группе. Их подсчитывают по формуле (5):
а = ркN / пк; к = 1,…, G, (5).
где рк — заданный вклад kй группы признаков в некоторую меру сходства ожидаемого потомства и заданного идеала; N — общее число признаков; пк — число признаков в k-й группе; G — число групп.
Общим недостатком методов, оценивающих близость к идеальному сорту средних значений признаков ожидаемого потомства, является отсутствие учета генетического разнообразия популяции потомства. Один вариант скрещивания может давать большую близость средних к идеалу, но иметь низкое генетическое разнообразие популяции потомства по изучаемым признакам, а второй — дальше от идеала по средним, но с гораздо большей генотипической изменчивостью. Во второй популяции потомства больше вероятность отобрать формы, близкие к идеальному сорту. Поэтому генетическое разнообразие потомства также оценивать и учитывать при подборе родительских форм.
Комбинационная способность и методы ее определения.
Общую комбинационную способность (ОКС), как правило, устанавливают на основе топкроссных испытаний. Для определения специфической комбинационной способности (СКС) необходимы диаллельные скрещивания, которые включают в себя все возможные комбинации скрещивания между серией сортов и линий. Соответствующие методы определения комбинационной способности разработал Б. Гриффинг, предложивший четыре метода анализа диаллельного скрещивания:
- 1. Изучают родительские формы, F1 — гибриды прямых и обратных скрещиваний всего m2 генотипов.
- 2. Изучают родительские формы и F1 полученные в результате прямых скрещиваний, — всего m (m+1)/2 генотипов.
- 3. Изучают только прямые и обратные гибриды F1 — всего m (m-1) генотипов.
- 4. Изучают только прямые гибриды F1 — всего m (m-1)/2 генотипов.
Для анализа комбинационной способности в целях гетерозисной селекции, как правило, применяют метод IV.
Топкроссы применяют для определения общей и специфической комбинационной способности при селекции на гетерозис. Для этого линии и сорта скрещивают с одним или несколькими сортами — анализаторами (тестерами). При этом в селекции самоопыляющихся растений большое значение имеет определение эффектов общей комбинационной способности, по величине которых можно судить о донорских свойствах скрещиваемых форм. Поликросс требует определенной подготовительной работы и планирования. Однако при использовании поликросса не требуется проводить оценку комбинационной способности родителей. Применяется главным образом к тем перекрестноскрещивающимся культурам, у которых получение семян от контралируемых скрещиваний затруднено. Результаты поликроссного испытания указывают на сорта, имеющие повышенную комбинационную способность, которые затем рекомендуют в состав синтетической популяции. По методам Гриффинга возможны два варианта оценки исходного материала. Вариант I (модель I) применяют, когда родительские формы для исследования отбирают специально и необходимо оценить их комбинационную способность. Формы одновременно являются тестерами, с помощью которых выявляют комбинации скрещиваний. Вариант II (модель II) используют, когда родительские формы отобраны случайно из популяции, которые необходимо оценить (тестером служит исследуемая родительская популяция). Исходные линии случайно отобраны из популяции, полученной длительным самоопылением (без отбора) генотипов исследуемой популяции. В данном случае представляют интерес не сами пара — метры отдельных родительских линий, а компоненты генотипической и дисперсии признака в исходной популяции. Значение признака для гибридной комбинации между i-м jм родительским сортом в k-м повторении можно выразить в биометрической модели следующий вид уравнения (6):
xijk=µ+gi+gj+sij+ri+ek (6).
где хijk — величина признака у гибрида F1 между i-й и j-й родительскими линиями в k-м повторении; µ- среднее значение признака в наборе линий и гибридов F1, gi, gj эффект общей комбинационной способности i-й и j-и родительских линии; sij — эффект специфической комбинационной способноcти; ri — реципрокный эффект при скрещиваний и j-й линий; ек — эффект, обусловленный случайной ошибкой в k-м повторении.
Существенность отношения определяемая по Р — критерию Фишера, указывает, что между исследуемыми гибридами (или гибридами и линиями) имеются различия по величине признака, случайные ошибки опыта. Если различия не значимы, дальнейший анализ комбинационной способности не проводят. Оценки и их разности, необходимые для дальнейшего анализа, несут случайные ошибки, которые характеризуются дисперсиями ошибок (var) (7):
(7).
Третий метод Гриффинга. Оценка одного из вкладов в дисперсию СКС меньше нуля. Это объясняется неучтенными ошибками опытов и, возможно, неполным соответствием данных стандартным предположениям дисперсионного анализа.
Теоретическая основа метода Хеймана. Данный метод базируется на предположении, что количественный признак детерминирован k полиморфными локусами, точнее блоками сцепленных полигенов. Каждый локус в анализируемом наборе из родительских линий может иметь два аллельных состояния. Вклад этих генных различий в оценки статистических параметров, сред — них, дисперсий, ковариаций будут следующими (без учёта ошибок опытов). Дисперсия величин признака у родительских линий (8):
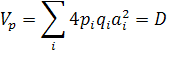
где? — сумма по всем k полиморфным локусам; D — изменчивость аддитивных эффектов аi у родителей с учетом частот pi и qi.
Метод Хеймана можно рассмотреть и как генетический анализ популяции. Из уравнении следует, что при выполнении требований на Э влияют только аддитивные, а на Н1 и Н2 — доминантные эффекты. Отношение характеризует степень доминирования в экспериментальном материале, среднюю по всем полиморфным локусам.
Отношения позволяют сделать выводы относительно генетической детерминации количественного признака у изученных родительских форм. В этом состоит так называемый параметрический вариант метода Хеймана. Используется также регрессионный анализ Хеймана. Метод Хеймана обладает определенной робастностью — устойчивостью к неполному удовлетворению шести требований. Вопрос о проверке выполнения требований и о пределах устойчивости выводов до настоящего времени решен не полностью. Сравнительно низкие требования к объёму и структуре эксперимента позволяют методу Хеймана оставаться популярным в селекционно-генетических исследованиях растений.