Двухмембранные оргаиеллы.
Физиология растений в 2 т. Том 1

Хлоропласта обладают собственной кольцевой ДНК (плДНК) и набором ферментов для синтеза белков. Однако собственная ДНК пластид не может полностью кодировать все специфические для пластиды белки. Гены для более 90% этих белков находятся в ядре (см. гл. 2). Установлено, что ДНК пластид содержит около 100 генов, которые делят на две группы: 1) гены, обслуживающие процессы транскрипции и трансляции… Читать ещё >
Двухмембранные оргаиеллы. Физиология растений в 2 т. Том 1 (реферат, курсовая, диплом, контрольная)
Пластиды (от греч. plastos — изменчивый, пластичный) есть только у растений. Каждая пластида окружена двойной мембраной — наружной и внутренней. Существует три основных типа пластид: хлоропласты, хромопласты и лейкопласты. Набор пластид в данной клетке зависит от ее дифференцировки. Из пластид наибольшее значение имеют хлоропласты, так как в них идет фотосинтез. Все пластиды имеют собственный геном и белоксинтезирующую систему (ферменты, рибосомы и пр.).
Хлоропласт (от греч. chloros — желто-зеленый) высшего растения похож на линзу, диаметр которой 5—8 мкм, а толщина — около 1 мкм. Наружная мембрана хлоропласта отделяет его от цитозоля.
Она проницаема для большинства органических и неорганических молекул. Наружная мембрана содержит специальные белки-переносчики (см. параграф 1.5), через которые в хлоропласт поступают пептиды из цитозоля.
Внутренняя мембрана хлоропласта обладает высокой избирательной проницаемостью для метаболитов. Она проницаема для маленьких незаряженных молекул, например для 02 и С02, а также для недиссоциированных низкомолекулярных монокарбоновых кислот. Большинство метаболитов (белки, липиды, органические кислоты и углеводы) может проникать через внутреннюю мембрану в строму только с помощью переносчиков.
Таким образом, мембраны, окружающие хлоропласт, контролируют обмен веществами между ним и цитозолем.
Кроме того, внутренняя мембрана является местом синтеза липидов и участвует в формировании гран и тилакоидов стромы. Тилакоид граны представляет собой плоский мешочек (от греч. thylacoides — мешковидный), ограниченный мембраной. Эти тилакоиды уложены в стопки — граны (рис. 1.11). В одной гране может находиться 10—30 тилакоидов, а в хлоропласте — до 100—150 гран. Тилакоиды стромы длинные, расположены параллельно длинной оси хлоропласта и соединяют граны друг с другом. Все пространство между гранами заполнено бесцветным раствором — стромой. Пространство внутри тилакоида называется внутритилакоидным пространством, или люменом (от англ, lumen — полость, просвет). В тилакоидах находятся пигменты и ферменты, принимающие участие в световой фазе фотосинтеза (см. гл. 4). В строме содержатся ферменты, участвующие в восстановлении С02.
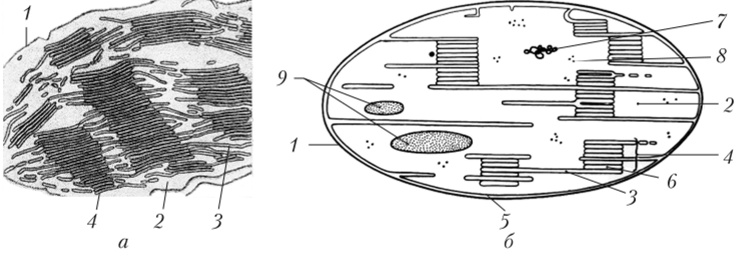
Рис. 1.11. Гранальный хлоропласт1:
а — фотография; б — схема; 1 — наружная мембрана; 2 — строма;
- 3 — тилакоид стромы; 4 — грана; 5 — внутренняя мембрана;
- 6 — тилакоид фаны; 7 — ДНК; 8 — рибосома; 9 — крахмальное зерно
Строма хлоропласта — водный раствор, содержащий водорастворимые соединения, неорганические ионы, ферменты, нужные для ассимиляции углерода и синтеза пигментов и липидов собственных мембран. В строме содержится своя специфическая кольцевая ДЫК (илДНК), которая отличается от ядерной и передается, но наследству через пропластиды яйцеклетки, а также РНК, рибосомы, ферменты для синтеза белков.
Строма содержит крахмальные зерна, образующиеся в хлоропластах днем, а также пластоглобулы, состоящие из липидов и пластохинонов. Особенно много пластоглобул содержится осенью в период осеннего листопада.
Хлоропласты, имеющие такое строение, получили название грапалъных (см. рис. 1.11). Однако у ряда растений (кукуруза, сахарный тростник, просо и многие другие) есть еще хлоропласты без гран, называемые агранальными.
Итак, хлоропласт можно разделить на три компартмента:
- 1) межмембранное пространство — между наружной и внутренней мембранами;
- 2) пространство между внутренней мембраной и мембранами тилакоидов, здесь находится строма;
- 3) пространство, окруженное гилакоидной мембраной, — внутритилакоидный люмен.
В строении хлоропласта мы можем наблюдать один из принципов построения живой материи — иерархию: так же, как организм состоит из органов, орган — из клеток, а клетка — из органелл, хлоропласт состоит из стромы и тилакоидов, тилакоиды образуются из мембран и сами могут образовывать граны.
В хлоропластах находятся все фотосинтетические пигменты (5—10% от его сухой массы). Кроме того, хлоропласт содержит 35—55% белков, 30—50% липидов, 2—3% РНК, 0,5% — ДНК, а также 5% золы, основными компонентами которой являются калий, фосфор, железо, медь, магний, цинк.
В хлоропластах происходит не только превращение световой энергии в химическую и ассимиляция углерода, но и ассимиляция фосфора, а также восстановление нитратов и серы. Кислород, находящийся в атмосфере Земли и необходимый для дыхания организмов и создания озонового слоя, образуется почти исключительно в хлоропластах при фотосинтезе.
Хлоропласты могут двигаться вместе с цитозолем (пассивно) или независимо от него. Их самостоятельные движения зависят от света. При слабом освещении хлоропласты располагаются возле освещенных передних и задних стенок и обращены к свету своей широкой стороной, что помогает лучшему поглощению квантов света. При сильном освещении хлоропласты передвигаются к боковым клеточным стенкам и поворачиваются к свету узкой стороной, что защищает их и хлорофилл от избытка света.
Гранальные хлоропласты могут делиться, но чаще они образуются из пропластид (рис. 1.12). Пропластида окружена двойной мембраной. Из внутренней мембраны пропластиды сначала возникает проламеллярпое тело. Затем, если света достаточно, из него формируются тилакоиды и граны. В мембраны тилакоидов встраиваются вновь синтезированные молекулы пигментов. Одновременно с ростом клеток растут и хлоропласты. Предполагают, что агранальные хлоропласты возникают в процессе формирования листовой пластинки из гранальных хлоропластов, так как сначала они тоже имеют граны.

Рис. 1.12. Образование хлоропласта1:
слева — развитие хлоропласта на свету (нормальное образование тилакоидов гран и тилакоидов стромы); справа — развитие хлоропласта в темноте (образуется проламеллярное тело) (Мюлетадер и Фрей-Вислинг, 1959)
Хлоропласта обладают собственной кольцевой ДНК (плДНК) и набором ферментов для синтеза белков. Однако собственная ДНК пластид не может полностью кодировать все специфические для пластиды белки. Гены для более 90% этих белков находятся в ядре (см. гл. 2). Установлено, что ДНК пластид содержит около 100 генов, которые делят на две группы: 1) гены, обслуживающие процессы транскрипции и трансляции белков пластид; их называют генами домашнего хозяйства; 2) гены белков, обеспечивающих работу пластид, прежде всего фотосинтез (см. гл. 2). Рибосомы, входящие также в белоксинтезирующую систему, либо свободно расположены в строме, либо прикреплены к поверхности тилакоидных мембран.
В клетке в среднем находится до 50 хлоропластов. В старой клетке хлоропласта разрушаются.
Хромопласты (от греч. chroma — цвет) но размеру меньше хлоропластов, форма их различна: шаровидные, нитевидные, изогнутые. Хромопласты содержат красные, желтые, оранжевые пигменты из группы каротиноидов, молекулы пигментов встроены в мембраны. Мембраны хромопластов состоят в основном из липидов. Внутренняя система мембран у них отсутствует. В строме много рибосом. Функция хромопластов до конца не изучена.
Хромопласты часто возникают из хлоропластов, одновременно при этом разрушаются рибосомы, а пластидные ДНК инактивируются. Хромопласты могут размножаться делением, могут образовываться из пронластид и обратно превращаться в хлоропласты, например в том случае, если корнеплод моркови попадает на свет.
Лейкопласты (от греч. leukos — белый) не содержат пигментов и характерны для прекративших рост клеток. По размеру они значительно меньше хлоропластов и не имеют строго определенной формы. Внутренняя мембрана у них развита плохо. В строме имеются молекулы ДНК, рибосомы.
Главная функция лейкопластов — синтез и накопление запасных веществ, в первую очередь крахмала, иногда — белков, редко — масел, в них также синтезируются изонреноиды (см. параграф 1.3).
В зависимости от химической природы запасаемого вещества лейкопласты делятся на амило-, протеинои элайопласты. Амилопласты содержат крахмал и встречаются в запасающих органах — корнях, семенах, клубнях; элайопласты содержат масла в многочисленных пластоглобулах, про те ино пласты — крупные белковые кристаллы. У некоторых растений в лейкопластах клеток корня происходит промежуточная реакция ассимиляции нитратов — восстановление нитритов до аммония (см. параграф 6.4). Существуют лейкопласты, в которых синтезируются изонреноиды.
В листьях растений, выращенных в темноте, образуются этиопласты (от фр. etioler — бледнеть, чахнуть). Они формируются из пропластид в темноте и считаются промежуточной стадией образования хлоропластов. Этиопласты не имеют хлорофилла и окрашены каротиноидами в бледножелтый цвет. Если зеленые части растения попадают в темноту надолго, то сначала разрушаются тилакоиды стромы, а потом гран, и из липидов разрушенных мембран возникает проламеллярное тело, а хлоропласта превращаются в этиопласты. В этиопластах содержится много протохлорофиллида (см. параграф 4.3). На свету этиопласты превращаются в хлоропласта, а из протохлорофиллида синтезируется хлорофилл.
Как уже было сказано, все пластиды (хлоро-, хромо-, лейко-, амило-, этиопласты) образуются из пропластид, они также могут размножаться делением и превращаться друг в друга. Чаще делятся пропластиды, этиопласты и молодые хлоропласта. Скорость деления иропластид в меристемах коррелирует со скоростью деления клеток, поэтому количество пластид в материнских и дочерних клетках сначала одинаково. Однако деление пластид необязательно связано с делением клетки, оно может происходить, например, в клетке в фазе растяжения (см. параграф 7.1).
Пропластиды являются предшественниками всех пластид и содержатся в основном в меристематических клетках, около 20 в каждой клетке. Это.
небольшие органеллы диаметром 1 — 1,5 мкм, окруженные двойной мембраной. В пропластиде есть кольцевые ДНК.
В пластидах происходит восстановление нитрита и сульфата, синтезируются все жирные кислоты, пурины, пиримидины, большинство аминокислот, изопреноиды (см. параграф 6.4). В зеленых клетках жирные кислоты синтезируются исключительно в хлоропластах, в незеленых — в хромо-, лейкопластах или в пропластидах. Часть жирных кислот используется для образования мембранных липидов пластид, часть — транспортируется в цитозоль.
Митохондрии — это органеллы, в которых происходит большая часть реакций дыхания. Освобождаемая при этом энергия аккумулируется в образующихся молекулах АТФ и используется для работы клетки, поэтому митохондрии часто называют энергетическими станциями клетки.
Митохондрия (от греч. mitos — нить и chondros — зернышко) обычно имеет короткую овальную или удлиненную форму (рис. 1.13). Длина митохондрии 4—7 мкм, а диаметр — 0,5—2 мкм. В состав этой органеллы входят в основном белки (60—65%) и липиды (около 30%), а также нуклеиновые кислоты (ДНК — 0,5% и РНК — 1%).

Рис. 1.13. Митохондрия:
1 — наружная мембрана; 2 — внутренняя мембрана; 3 — матрикс; 4 — криста; 5 — АТФ-сомы на кристе внутренней мембраны.
Митохондрия окружена двойной мембраной. Между мембранами находится пространство, называемое межмембранным. Внутреннее пространство митохондрии заполняет бесцветный раствор — матрикс. В матриксе находятся разные ферменты, в том числе участвующие в дыхании.
Внутренняя мембрана образует выросты — кристы, расположенные перпендикулярно продольной оси органеллы. Количество крист зависит от вида организма, тина ткани, возраста клетки. Кристы увеличивают поверхность внутренней мембраны, в которой расположены цепь транспорта электронов (дыхательная цепь) и молекулы аденозинтрифосфатазы (АТФазы), катализирующей образование АТФ из АДФ и неорганического фосфата (см. параграф 1.2).
Как и у хлоропластов, наружная мембрана имеет высокую проницаемость. Она содержит белки порины и проницаема для небольших моле-
кул размером 4—6 кДа. Внутренняя мембрана практически непроницаема не только для молекул и ионов, но даже для протонов, иначе было бы невозможно накопление электрических зарядов, необходимое для синтеза АТФ (см. параграф 5.2). Вещества транспортируются через эту мембрану с помощью белков-переносчиков или через специальные каналы.
Кроме дыхания, в митохондриях происходит превращение глицина в серин, т. е. часть гликолатного цикла фотосинтеза (см. гл. 4). Митохондрии также участвуют в запрограммированной смерти клеток (см. гл. 8) и ионном обмене, например в них находится запас ионов кальция.
Число митохондрий в клетках разных тканей различно и зависит от метаболической активности клетки. Небольшие клетки кончика корня содержат по 200 митохондрий, а клетки зрелого корня — по 2000. В клеткесиутнице ситовидной трубки до 20% от объема се цитоплазмы приходится на долю митохондрий.
Образуются митохондрии из инициальных частиц, существующих в меристематических клетках и представляющих собой пузырьки, окруженные двойной мембраной (рис. 1.14). Инициальная частица увеличивается в размере, а ее внутренняя мембрана начинает образовывать кристы. Так из инициальной частицы формируется промитохондрия. Последняя продолжает расти, число крист в ней увеличивается, и она превращается в митохондрию. Митохондрии могут размножаться делением с помощью перетяжки или почкованием. При почковании мембраны митохондрии выпячиваются, образуется вырост (почка), который затем отделяется. Часто митохондрии образуют комплексы с хлоропластами, пероксисомами, ядром, эндоплазматической сетью.

Рис. 1.14. Образование митохондрии:
1 — инициальная частица; 2 — нромитохондрия; 3 — митохондрия
Как и хлоропласта, митохондрии обладают собственной системой синтеза белка, состоящей из ДНК и 705-рибосом. Кроме крупных кольцевых молекул ДНК, митохондрии содержат некоторое количество кольцевых и линейных молекул меньшего размера. Однако геном митохондрий кодирует лишь незначительную часть митохондриальных белков, а большая часть кодируется ядерным геномом.
Таким образом, в клетке имеется три относительно самостоятельных генома — ядерный (хромосомный), пластидный и митохондриальный.