Транспирация как саморегулируемый процесс

Существует ли связь между устьичной и внеустьичной регуляцией транспирации? Рассмотрим график зависимости интенсивности транспирации от ширины устьичной щели (рис. 3.21). Кривая показывает, что, когда устьица только начинают открываться, интенсивность транспирации растет почти линейно. Однако когда устьица широко открыты, дальнейшее увеличение щели не влияет заметно на интенсивность транспирации… Читать ещё >
Транспирация как саморегулируемый процесс (реферат, курсовая, диплом, контрольная)
Для нормального роста растений высокая интенсивность транспирации не нужна. Во влажном тропическом климате или в теплицах с повышенной влажностью воздуха, где транспирация снижена до минимума, растения достигают больших размеров. Сами растения выработали в процессе эволюции различные приспособления для сокращения транспирации (см. параграф 9.1). Однако уменьшать транспирацию очень сильно нельзя, так как она является верхним концевым двигателем водного тока. Из этого противоречия природа нашла выход, создав осциллирующие механизмы, о которых уже было сказано выше.
Различают два типа регуляции транспирации: устьичпый и виеустьичный. Устьичная регуляция осуществляется с помощью открывания и закрывания устьиц. Закрывание устьиц наполовину мало влияет на интенсивность транспирации, что вытекает из закона Стефана (см. параграф 3.4). Полное их закрывание сокращает транспирацию примерно на 90%.
Внеустьичная регуляция транспирации — это, прежде всего, уменьшение испарения воды в межклетники, которое осуществляется с помощью нескольких механизмов. Первый механизм связан с обезвоживанием стенок клеток мезофилла, с поверхности которых идет испарение. Этот механизм назвали механизмом начинающегося подсушивания. Если в лист приходит меньше воды, то клеточные стенки хлоренхимы начинают подсыхать. Если воды мало, водные мениски в капиллярах между фибриллами целлюлозы становятся вогнутыми, что увеличивает силы поверхностного натяжения, и испарение идет медленнее.
Второй механизм связан со способностью цитоплазмы связывать воду. На его существование указывает тот факт, что при одной и той же степени открытости устьиц интенсивность транспирации может сильно меняться. Крахмал может превратиться в глюкозо-1-фосфат не только в замыкающих клетках устьиц, но и в клетках хлоренхимы. В результате произойдет уменьшение водного потенциала этих клеток, а значит, и транспирации.
Может изменяться толщина кутикулы, покрывающей листья. Увеличение ее толщины уменьшает транспирацию с поверхности листьев. Наблюдения с помощью электронного микроскопа показали, что в кутикуле могут образовываться трещины, что увеличивает кутикулярную транспирацию. Кроме того, кутикула обладает интересным свойством: при подсыхании наружных стенок клеток эпидермы слои кутикулы плотнее придвигаются друг к другу, и испарение воды уменьшается. При увеличении оводненности эпидермы кутикула набухает, разрыхляется, и кутикулярпая транспирация увеличивается. Таким образом, скорость кутикулярной транспирации может регулироваться оводнениостыо листовой пластинки. По ночам, при более сильном набухании кутикулы, кутикулярная транспирация идет быстрее, чем днем. Эти механизмы уменьшения интенсивности транспирации выгодны тем, что не влияют на поглощение углекислого газа.
Существует ли связь между устьичной и внеустьичной регуляцией транспирации? Рассмотрим график зависимости интенсивности транспирации от ширины устьичной щели (рис. 3.21). Кривая показывает, что, когда устьица только начинают открываться, интенсивность транспирации растет почти линейно. Однако когда устьица широко открыты, дальнейшее увеличение щели не влияет заметно на интенсивность транспирации. Следовательно, при максимально открытых устьицах скорость транспирации регулирует в основном внеустьичный механизм. При закрытых устьицах уменьшение кутикулярной транспирации происходит за счет внеустьичной регуляции.
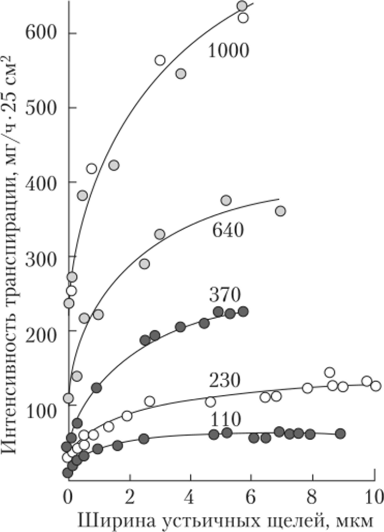
Рис. 3.21. Зависимость интенсивности транспирации от ширины устьичной щели1:
числа на кривых — скорость испарения со свободной водной поверхности
У некоторых растений существуют свои специфические механизмы внеустьичной регуляции транспирации. Например, у злаков, когда транспирация превышает поступление воды, листовые пластинки свертываются в трубку вдоль средней жилки. Свертывание уменьшает интенсивность транспирации на 10—20%. У банана при сильном водном дефиците обе половинки листа складываются. В результате между ними повышается относительная влажность воздуха, и транспирация уменьшается. Складывание листьев объясняется изменением тургорного давления в гиподерме. Некоторые исследователи считают, что по величине угла между половинками листа можно даже судить о степени водного дефицита.[1]
Поскольку количество воды, теряемой растением, зависит не только от интенсивности транспирации, но и от общей испаряющей поверхности побега или всего растения, то при недостатке воды растение может терять листья. Например, у сахарного тростника в условиях засухи на побеге может остаться только один лист. Одновременно недостаточное поступление воды вызывает торможение растяжения клеток, в результате задерживается рост новых листьев, образуются более мелкие листья, т. е. сокращается транспирирующая поверхность.
Листопад у деревьев умеренной зоны — это приспособление к зимней засухе. Зимой из-за низкой температуры почвы корни плохо дышат, поэтому плохо поглощают соли, и в результате вода медленно поступает в растение. Однако листопадные деревья встречаются и в тропиках, и в пустынях. Эти деревья теряют листья в начале сухого сезона.
Участие разных механизмов в регуляции транспирации у растений разных видов неодинаково. У оливковых деревьев главную роль в регуляции играет устьичный механизм, у хлопчатника преобладает внеустьичный. У растений умеренной зоны впеустьичное регулирование транспирации выражено меньше — без закрывания устьиц транспирация может быть уменьшена на 30%.
У гигрофитов нет приспособлений, ограничивающих расход воды. В их листьях мало устьиц, которые всегда открыты, а кутикула развита слабо, поэтому кутикулярная транспирация велика. Избыток воды выделяется наружу с помощью гидатод.
У полностью погруженных в воду гидрофитов транспирация отсутствует. Основной механизм регулирования водного обмена — ограничение поступления воды, которая поступает через всю поверхность.
Итак, регуляция водного обмена происходит постоянно, особенно в жаркие дни. Если днем транспирация идет быстро, возникает водный дефицит. Закрываются устьица, транспирация уменьшается, а корни одновременно поглощают больше солей. Это вызывает уменьшение водного потенциала, поэтому скорость поступления воды ночью увеличивается. Поступление воды ликвидирует дневной дефицит воды. Утром транспирация усиливается и постепенно опять начинает превышать свое поступление.
- [1] Цит. по: Сабинин Д. А. Физиологические основы питания растений.