Ионные каналы.
Нервная система: анатомия, физиология, нейрофармакология

Один из сильнейших ядов содержится в тканях рыбы-иглобрюха, называемой японцами рыбой фугу. У этих тропических рыб особенно ядовиты печень и яичники. Несмотря на это, иглобрюх является деликатесом японской кухни. Яд из тканей этих рыб называется тетродотоксином, и достаточно попадания в организм человека всего лишь 0,5 мг токсина, чтобы он погиб. Для сравнения: знаменитый цианистый калий… Читать ещё >
Ионные каналы. Нервная система: анатомия, физиология, нейрофармакология (реферат, курсовая, диплом, контрольная)
Таким образом, в момент нанесения на возбудимую клетку внешнего раздражителя в нее в течение очень короткого времени входят ионы натрия, и мембрана клетки деполяризуется настолько, что наблюдается овершут, но в следующее мгновение резко возрастает ток К+ из клетки, в результате чего потенциал на мембране возвращается к уровню покоя: -70 мВ. Напомним, что в покое мембрана возбудимых клеток вполне проницаема для К+, а для Na+ является практически непреодолимым препятствием.
Так каким же образом обеспечиваются токи ионов через мембрану в момент генерации ПД? Оказывается, в клеточной мембране возбудимых клеток располагаются особые белковые молекулы, образующие ионные каналы. Восходящая фаза ПД реализуется за счет работы потенциалзависимых натриевых каналов. Когда клетка находится в состоянии покоя, этот канал закрыт (рис. 8.8).
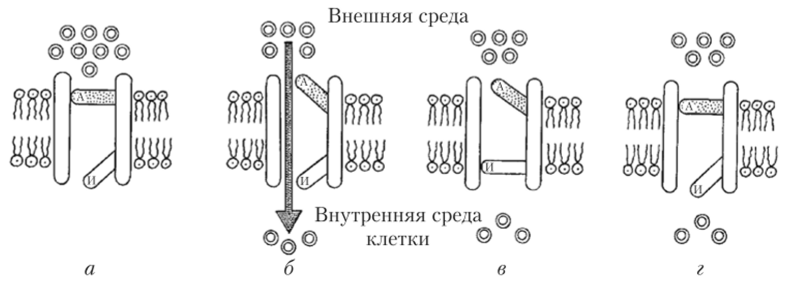
Рис. 8.8. Схема работы потенциалзависимого натриевого канала:
а — канал закрыт; 6 — канал открыт; в — канал инактивирован; г — канал возвращен в исходное состояние и готов к работе; А — активационная створка;
И — инактивационная створка Если же клетку раздражают каким-либо внешним стимулом, то ПП на мембране начинает сдвигаться от -70 мВ к -50 мВ. Для большинства нервных клеток достаточно сдвига ПП на 20 мВ (т.е. до -50 мВ) для того, чтобы пространственная конфигурация белковой молекулы канала изменилась, образуя проход, по которому в клетку устремляются по градиенту концентрации ионы натрия. Происходит это из-за того, что часть молекулы канала (которую назвали активационной створкой канала) сдвигается, и открывается канал для ионов натрия. Однако в следующее мгновение другая часть канальной молекулы белка (названная инактивационной створкой канала) перекрывает движение ионов через канал (см. рис. 8.8).
Натриевый канал находится в открытом состоянии всего 1 мс, но за это время через него в клетку успевает пройти около 6 тыс. ионов Na+. Конечно, работа одного канала не может заметно деполяризовать клетку, но натриевых каналов на мембране возбудимых клеток очень много — около 100 на площади в 1 мкм2. После срабатывания канал за несколько миллисекунд возвращается в исходное состояние и может открыться снова, если клетка подвергнется раздражению (см. рис. 8.8). Но для того, чтобы Na+-Kana.T перешел из состояния, изображенного на рис. 8.8, в, в состояние, изображенное на рис. 8.8, а, необходимо, чтобы потенциал на мембране достиг ПП. Восстановление ГГП на мембране осуществляется благодаря тому, что сразу же после закрытия натриевого канала через свои особые каналы из клетки выходят положительно заряженные ионы К+. Калиевые каналы действуют по тем же принципам, что и натриевые, однако у них есть только одна створка, которая открывает и закрывает путь ионам калия из клетки.
Ионные каналы обладают избирательностью: пропускают только «свои» ионы. Однако бывают и исключения. Например, ионы лития (Li+) очень хорошо проходят через натриевые каналы внутрь возбудимых клеток (даже немного лучше, чем сам натрий). Но лития в организме человека практически нет. Литий применяется для изменения работы мозга в случае некоторых тяжелых психических расстройств.
В настоящее время открыто несколько видов натриевых и калиевых каналов. Они различаются величиной порога открывания, скоростью открывания и закрывания, пропускной способностью для ионов, механизмом открывания.
Существуют также особые малоизбирательные каналы для анионов и катионов и, наоборот, очень избирательные каналы для Са2+ и С1″.
Сразу после того, как активационная створка пропускает в клетку порцию натрия, канал снова активировать невозможно (см. рис. 8.8), так как необходимо сначала вернуть в исходное состояние инактивационную створку. В этом состоянии канал называют абсолютно рефрактерным, т. е. неспособным к работе. По длительности абсолютная рефрактерность равна приблизительно 1 мс. Наличие абсолютной рефрактерности не позволяет отдельным ПД накладываться друг на друга и ограничивает максимальную частоту разрядов нервных клеток величиной 500—700 Гц. Вслед за стадией абсолютной рефрактерное™ наступает относительная рефрактерность: лишь часть Ыа+-капалов вернулась в состояние готовности, еще не закрыты все потенциалзависимые К+-каналы. В таких условиях запуск ПД возможен, но только под действием более мощного стимула.
Нормальная работа ионных каналов очень важна для организма. Ведь нарушение их работы приводит к очень быстрому развитию нарушений функционирования ЦНС, судорогам, параличам и т.и. — в зависимости от того, какой вид каналов дезорганизован.
Природа очень далеко зашла по пути создания блокаторов ионных каналов, которыми по справедливости вооружила и хищников, и тех, кто от этих хищников обороняется.
Один из сильнейших ядов содержится в тканях рыбы-иглобрюха, называемой японцами рыбой фугу. У этих тропических рыб особенно ядовиты печень и яичники. Несмотря на это, иглобрюх является деликатесом японской кухни. Яд из тканей этих рыб называется тетродотоксином, и достаточно попадания в организм человека всего лишь 0,5 мг токсина, чтобы он погиб. Для сравнения: знаменитый цианистый калий приблизительно в 100 раз менее ядовит. Тетродотоксин действует с наружной стороны натриевого канала, блокируя вход в него натрия. В результате генерация ПД становится невозможной, что приводит к расслаблению мускулатуры и параличу дыхания.
Еще более страшный яд содержится в коже внешне совершенно безобидных лягушек из рода листолазов, живущих в Южной Америке. Этот яд, называемый батрахотоксином, ядовитее цианистого калия в 500 раз! Одна маленькая лягушка носит в коже столько яда, что им можно погубить до 350 человек. Батрахотоксин блокирует натриевый канал в момент его открытия, не давая створкам захлопнуться, после чего он не может закрыться, и натрий, поступая в клетку без ограничений, приводит к ее перевозбуждению. В результате, как правило, первыми нс выдерживают клетки сердечной мышцы, и сердце перестает биться.
Похожим эффектом обладают яды скорпионов, поддерживающие натриевые каналы в открытом положении. Блокаторы натриевых каналов вырабатывают и водоросли (сакситоксин), и высшие растения (аконитин).
Еще одна группа веществ, оказывающих свой лечебный эффект через воздействие на натриевые каналы, — местные анестетики (новокаин, лидокаин, прокаин и многие другие). Молекулы этих веществ проникают в канал изнутри клетки, снижая натриевые токи и блокируя проведение электрических сигналов, но тонким безмякотным нервным волокнам. Как раз по таким волокнам передастся в мозг информация о болевых воздействиях. Таким образом, местные анестетики на время блокируют болевые ощущения.
Блокаторов К+-каналов в природе мало, зато создан синтетический блокатор — тетраэтиламмоний, «затыкающий» канал изнутри клетки.
В настоящее время известно множество типов Na+— и К+-каналов, характерных для тех или иных клеток организма. Эти каналы отличаются друг от друга условиями, в которых они открываются и закрываются, пропускной способностью, скоростью открывания и закрывания и т. п. Кроме того, найдены каналы для С1~, Са2+ и т. п.
Значительную роль во всех возбудимых тканях играют кальциевые токи, идущие по градиенту концентрации в клетку. Существуют специфические потенциалзависимые каналы, через которые Са2+ входит в клетку, выполняя при этом две функции: как положительно заряженный ион — деполяризует мембрану и как мощный внутриклеточный регуляторный фактор — меняет работу всех систем клетки.
Очень важны кальциевые токи в процессах возбуждения мышц и передачи информации от клетки к клетке. В эволюции каналы для кальция, по-видимому, возникли очень рано и Са2+-ток через мембрану древнее Ка+-тока. Каналы для кальция открываются относительно медленно, плотность их на мембране меньше, чем каналов для Na+, но зато Са2+-каналы не закрываются до конца, обеспечивая постоянный слабый вход этого иона в клетку. Блокаторами кальциевого тока через каналы являются многие двухвалентные ионы: кадмий, кобальт, марганец и др.
В заключение этого параграфа еще раз сравним работу ионных каналов и ионных насосов (Ыа+/К+-АТФ-аза). Задача обеих систем — транспорт ионов через мембрану возбудимой клетки, но на этом их сходство и заканчивается.
Сравнение пассивного (каналы) и активного (насосы) транспорта Na+ и К+ через клеточную мембрану.
Таблица 8.1
Характеристика. | Каналы. | Насос. |
Направление движения ионов. | По электрохимическому градиенту. | Против градиента. |
Источник энергии. | Градиент ионов. | Энергия АТФ. |
Характеристика. | Каналы. | Насос. |
Зависимость проводимости от потенциала на мембране. | Зависит. | Не зависит. |
Блокаторы транспорта. | Тетродотоксин. | Дигитонин, сапонин, строфантин. |
Влияние температуры. | Влияние малое. | Охлаждение тормозит транспорт. |
Скорость транспорта. | Во время восходящей фазы ПД — 10 s моль/см2 • с. | 6 • 10″ 11 моль/см2 • с. |
Влияние блокаторов энергетического обмена. | Не влияют. | Блокаторы прекращают работу насоса (цианиды, динитрофенол). |
Очевидно, что транспорт ионов через каналы и насосы — совершенно различные независимые процессы, однако активный транспорт (насосы) совершенно необходим для того, чтобы обеспечить поддержание разности содержания Na+ и К+ по разные стороны мембраны, что и определяет возможность возбудимой клетки удерживать ПП и генерировать ПД путем перемещения ионов через каналы.